의학박사학위논문 차세대염기서열분석기법을이용한 한국인의미토콘드리아 전체염기서열분석 Entire Mitochondrial DNA Sequencing on Massively Parallel Sequencing for the Korean population 2017 년 8 월
|
|
|
- 도한 반
- 5 years ago
- Views:
Transcription






1 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다. 저작권자로부터별도의허가를받으면이러한조건들은적용되지않습니다. 저작권법에따른이용자의권리는위의내용에의하여영향을받지않습니다. 이것은이용허락규약 (Legal Code) 을이해하기쉽게요약한것입니다. Disclaimer
2 의학박사학위논문 차세대염기서열분석기법을이용한 한국인의미토콘드리아 전체염기서열분석 Entire Mitochondrial DNA Sequencing on Massively Parallel Sequencing for the Korean population 2017 년 8 월 서울대학교대학원 의학과법의학전공 박소형
3 초 록 미토콘드리아의 디옥시리보핵산 (DNA) 염기서열분석은 유전적 다양성을파악할수있는중요한분석도구로, 인류의기원을알기위한 계통발생학분야에서뿐만아니라그유전적다양성을바탕으로하는 법의학의개체식별영역에서도유용한방법으로사용되어왔다. 종래의 미토콘드리아 DNA 염기서열분석기법은 실무적인 측면에서 비용 및 시간적인 효율성 때문에 조절부위 (the control region) 등 일부 염기서열에 국한하여 시행되어 왔었고, 따라서 그에 따른 유전적 다양성에대한정보도제한적일수밖에없었다. 인간유전체사업이후 방대한 유전학적 정보를 효율적으로 분석할 수 있는 새로운 기술의 필요성이 대두되면서 차세대염기서열분석기법이 개발되었다. 이러한 유전학의기술적발전은적은비용과시간으로도미토콘드리아의전체 염기서열분석이가능하게되어, 기존의방법으로는제한적이었던유전적 특성에 대해서도 손쉽게 접근할 수 있게 되었다. 그리하여 여러 인종에서나타나는미토콘드리아 DNA의유전적다양성에대한새로운 정보를알아내고자하는연구들이시도되기시작하였다. 또한법의학 개인식별측면에서도더많은추가적인정보를통해개인식별력을높일 수 있을 것으로 기대되었다. 한국인의 경우 기존의 대부분의 분석 자료들은주로조절부위, 암호부위일부등에국한된것이었으며그에 따른유전적다양성에대한정보도제한적인상황이다. 따라서이러한 연구변화의흐름에따라한국인의미토콘드리아 DNA 전체염기서열분석 및그에따른유전적다양성에대한연구가필요하다고보았다. 따라서 본연구를통해한국인의미토콘드리아 DNA 전체염기서열을분석하여 한국인에서나타나는유전적특성을파악하고이를바탕으로한국인에서 법의학적으로 개인식별력을 높일 수 있는지 알아보고자 하였다. 한국인에서 주로 나타나는 하플로그룹 (haplogroup, 조절부위 근거 ) 의 빈도를 고려하여 186명의 한국인 시료를 선별하여 차세대 기서열분석기법인 Ion Torrent Personal Genome Machine (PGM) 기술 (Thermo-Fisher사) 을 적용하여 미토콘드리아 DNA 전체 염기서열분석을시행하였다. 분석결과중국등동아시아민족과다른 대륙의민족과비교하여보았을때, 한국인에서나타나는각조절부위 및 암호부위별 변이의 구성과 분포, 이형세포질성의 분포 등은 전반적으로 다른 민족과 유사하였으나, 각 민족에서 공통적으로 i
4 나타나는주요변이와위치는중국과일본등동아시아민족안에서는거의유사하지만다른민족에서는다르게나타났다. 전체염기서열분석에따른하플로그룹의유형과세분화, 하플로타입 (haplotype) 에대한분석결과, 조절부위에근거하였을때의하플로그룹분류보다전체염기서열분석결과세분화되는양상을보여주었으며, 조절부위로결정되었던하플로그룹이전체염기서열분석결과다른하플로그룹으로재분류되었다. 이는중국등동아시아민족과다른대륙의민족에서도유사한양상을보였고, 특히한국인을포함하는동아시아인종의경우매크로하플로그룹 (macrohaplogroup) M(D, G, M) 이주요하플로그룹을이룬다는점에서, 전체염기서열분석정보는유전적다양성을파악하는데필수적이며, 조절부위등에국한된정보는유전적다양성을파악하는데제한적임을알수있었다. 이와함께한국인에서는개체수에비해하플로그룹개수가낮은편이지만하플로타입의개수는개체수에근접한점역시전체염기서열분석이개체식별력을높여줄수있음을보여주는것이었다. 이와함께동일한하플로그룹안에서서로다른변이즉개별변이 (private mutation) 의발견등은한국인에서잠재적인새로운하위하플로그룹의가능성을고려해볼수있는소견으로생각되었다. 한국인의미토콘드리아 DNA에서도이형세포질성이관찰되었으며그분포와양상은다른민족에서의연구결과와유사하였다. 본연구가앞으로한국인에서미토콘드리아 DNA의유전적다양성및질병연구에대한연구에견인차역할을할것으로기대된다. 주요어 : 법의학, 미토콘드리아, mtdna, 차세대염기서열분석, 한국인학번 : ii
5 목 차 서론 1 1. 미토콘드리아 DNA 미토콘드리아의기원 미토콘드리아의신원 미토콘드리아의유래 미토콘드리아의역사 미토콘드리아의유전시스템 미토콘드리아의생장과미토콘드리아 DNA 미토콘드리아 DNA 복제 미토콘드리아 DNA 변화 미토콘드리아 DNA 복구시스템 미토콘드리아 DNA 변화와복구시스템간의불균형및유전시스템에미치는영향 미토콘드리아 DNA의유전적특성 미토콘드리아 DNA 유전방식 : 모계유전 미토콘드리아 DNA 유전과이형세포질성 미토콘드리아 DNA의구조 미토콘드리아내 DNA의구조 미토콘드리아 DNA 가닥의구조와구성 세포핵미토콘드리아 DNA 염기서열 (numt) 법의학에서미토콘드리아 DNA의의의 법의학에서활용되는미토콘드리아 DNA의특성 미토콘드리아 DNA와세포핵 DNA 미토콘드리아 DNA 분석의장점 개체식별측면에서미토콘드리아 DNA의특징 미토콘드리아 DNA와계통발생학및법의학 미토콘드리아 DNA와인류의진화연구 미토콘드리아 DNA로인류계통발생나무구축 미토콘드리아 DNA DB 구축 미토콘드리아 DNA와하플로그룹 23 iii
6 2.3 미토콘드리아 DNA의법의학적활용의역사 미토콘드리아 DNA를이용한개인식별 미토콘드리아 DNA 이형세포질성과개인식별 한국에서미토콘드리아 DNA와개인식별 법의학에서미토콘드리아 DNA의최근연구동향 법의학에서 미토콘드리아 DNA에 대한 기존 연구의 한계 미토콘드리아 DNA 분석의한계 기존미토콘드리아 DNA 분석기법의한계 차세대염기서열분석기법의도입 새로운 DNA 분석기법의필요성대두 생거기법의기술적변화 : 피로시퀀싱 차세대염기서열분석기법의탄생 Ion Torrent PGM의개발 법의학에서차세대염기서열분석기법을 이용한미토콘드리아 DNA 연구 차세대염기서열분석기법을적용한미토콘드리아 DNA 염기서열분석기법의입증 다양한인종에서미토콘드리아 DNA 유전적다양성 연구 미토콘드리아 DNA 전체염기서열분석도구의변화 동아시아인종, 한국인에서미토콘드리아 DNA에대한연구 연구의필요성 36 목적 38 재료와방법 시료 미토콘드리아 DNA 추출과증폭 라이브러리준비와초병렬염기서열분석 40 iv
7 4. 자료분석 윤리적선언 41 연구결과 미토콘드리아 DNA 염기서열분석과 coverage 염기서열분석과 coverage 염기서열부위에따른 coverage 미토콘드리아 DNA 염기서열에서나타나변이 변이검출및불일치해석 보고되지않은변이검출오류 삽입-결실과관련된정렬오류 기타변이검출 동형세포질성변이 공통변이 개별변이 하플로그룹분류 하플로그룹 D 하플로그룹 A5a 하플로그룹 M 하플로그룹 F1b 하플로그룹 N9a2a 하플로그룹 G1a 하플로그룹 Y 하플로그룹 B 하플로그룹 M9;G;D 하플로타입 점이형세포질성 점이형세포질성의정의 점이형세포질성에서 coverage 점이형세포질성의분포와양상 하플로그룹 D 하플로그룹 A5a 60 v
8 하플로그룹 M 하플로그룹 F1b 하플로그룹 G1a 하플로그룹 B 하플로그룹 N9a2a 하플로그룹 M9;G;D 하플로그룹 Y 동일개체에서다수이형세포질성변이 염기서열부위와이형세포질성변이 하플로그룹을결정하는변이부위에형성된점이형세 포질성변이 세포핵미토콘드리아 DNA 염기서열 (numt) 과점이형세포 질성 66 고찰 차세대염기서열분석기법에의한미토콘드리아 DNA 전체염기 서열분석 염기서열부위에따른 coverage 변화 VCF의변이검출오류 한국인에서미토콘드리아 DNA 유전적특성 한국인에서변이의분포와양상 한국인에서공통변이양상 한국인에서하플로그룹의세분화양상 한국인에서하플로타입의양상 한국인에서개별변이양상 한국인에서미토콘드리아 DNA 점이형세포질성 한국인에서미토콘드리아 DNA 전체염기서열분석과개체식별 력 75 결론 77 참고문헌 78 vi
9 영문초록 (Abstract) 91 표 94 그림 122 부록 131 vii
10 표목차 [ 표 1] 186명의한국인에서하플로그룹분포구성 [ 표 2] 186명의한국인의인구학적특성. 95 [ 표 3] 미토콘드리아 DNA 전체염기서열분석을위한길잡이 [ 표 4] 변이검출형식보고서에서오검출된변이 [ 표 5] IGV를이용한오검출된변이검토 [ 표 6] 미토콘드리아 DNA 동형세포질성변이분포 [ 표 7] 하플로그룹에따른개별변이분포 [ 표 8] 한국인에서하플로그룹세분화양상 [ 표 9] 하플로그룹을결정하는주요변이 [ 표 10] 하플로타입의양상 [ 표 11] 점이형세포질성변이분석 [ 표 12] 점이형세포질성변이분포 [ 표 13] 염기서열분석에서 coverage 비교 114 [ 표 14] VCF 변이검출오류비교분석 115 [ 표 15] 동형세포질성변이의민족간비교분석 116 [ 표 16] 공통변이의민족간비교분석 [ 표 17] 하플로그룹세분화양상민족간비교분석 [ 표 18] 하플로그룹재분류비교분석 120 [ 표 19] 점이형세포질성비교분석 viii
11 그림목차 [ 그림 1] 미토콘드리아 DNA 구조 [ 그림 2] 미토콘드리아 DNA에따른인류의이동과분포 123 [ 그림 3] 인류계통발생나무. 124 [ 그림 4] 한국인에서주요하플로그룹 125 [ 그림 5] 동아시아인에서주요하플로그룹 126 [ 그림 6] Ion Torrent PGM 기법. 127 [ 그림 7] 한국인에서하플로타입양상및민족간비교분석 [ 그림 8] 전체염기서열분석결과하플로그룹 / 하플로타입변화 129 [ 그림 9] 전체염기서열분석결과하플로타입식별력증가 ix
12 서론 1. 미토콘드리아의 DNA 1.1. 미토콘드리아의기원 미토콘드리아의신원 미토콘드리아는세포의세포질안에있는작은구조물을말한다. 이구조물은이중막으로둘러싸여있는구조이다. 주요기능으로는세포내생명활동을수행하기위해에너지가필요하기때문에음식에서소화과정을통해얻어진포도당등의물질을산화하여 ATP 등의에너지를생산하는것이다. 이러한기능적특성뿐만아니라미토콘드리아의크기가작은세균과크기가유사하다는점, 세포의핵 DNA와는별개로독립적인원형의 DNA 및리보솜이있는점에착안하여. 미토콘드리아는세균에서유래하였을것이라는가설을제시하였다. 최초의원시세포는산소를이용한대사활동을할수없었다. 그리하여산소를이용한대사작용을할수있어서독립적으로생존이가능한세균들을포식하여생존에도움을받으려하였을것이다. 또한포식당한세균들은원시세포안에서생존하려하였을것이기에자기들을포식한원시세포와공생하기로하였을것이다. 그결과세균들은적대적인외부환경에서보호받을수있는안식처와영양분을공급받고, 대신원시세포에게에너지를공급하여생명활동을유지할수있도록하였을것이다. (Alberts B et al., 2008, Scheffler IE., 2008) 미토콘드리아의유래 미토콘드리아연구는 1800년대후반에들어서야시작되었다. 이는그 당시 탄생한 현미경과 조직 염색기술의 덕택으로 세포와 세포내 구조물을 직접 관찰함으로서 그들의 존재를 인지할 수 있었기 때문이기도하였다. Altman은 1890년 Elementarorganismen 이라는 책에서 세포질안에서 과립처럼 보이는 물질 ( 오늘날의 미토콘드리아 ) 을 bioplast 라명명하였고, 이들과립과같은물질은 숙주 세포의 세포질에 생존하는 세균과 유사한 집락을 형성하면서 1
13 자율적이고생존에중요한역할을할것이다 라고제시하였는데, 이는현대의미토콘드리아의유래에대한가설과매우유사함을알수있다. 1918년에들어서, 우리가미토콘드리아로알고있는세포내구조물에대한여러연구들을검토후종설이발표되었는데당시미토콘드리아는다양한이름으로설명되었다. blepharoblasts, chondriokonts, chondriomites, chondrioplasts, chondriosomes, chondriospheres, fila, fuchsinophilic granules, Korner, Fadenkorper, mitogel, parabasal bodies, plasmasomes, plastochondria, plastosomes, vermicules, sarcosomes, interstitial bodies, bioblasts 등으로언급되었다. 이러한여러용어들중오늘날의 mitochondria ( 미토콘드리아 ) 라는용어로정착된것은세포질에서관찰되는미토콘드리아의모양을묘사하던데에서유래하였다. 첫째현미경으로미토콘드리아를직접관찰하였을때과립처럼보인다고하여그리스어인 Chondros, 영어의 grain, 독일어의 Korn 등을이용하여묘사하였었다. 둘째염색기술개발덕택으로미토콘드리아를더상세하게묘사할수있었던점에서유래하였다. 1800년대후반에서부터핵과세포질, 세포내구조물을쉽게구별하여보여주는염색기법을고안하기위해부단히노력하였고그결과여러가지염색기법이개발되었다. 염기성염색시료와산성염색시료를이용하면조직학적으로다른색깔의염색으로세포와조직구조물을감별해줄수있다는발견을바탕으로 1896년오늘날보편적으로사용하는헤마톡실린-에오신염색기법이개발되었다. 세포구조물을염색하여구분하여관찰할수있게되면서, 일부세포에서는미토콘드리아가실처럼보이는구조물로관찰되었고, 이를그리스어의 mitos로서묘사하였다. 따라서실을뜻하는 mitos와과립또는미립자를뜻하는 Chondria가합성되어형성된미토콘드리아, mitochondria 라는용어가탄생하였다. (Andrews RM, et al., 1999, Rosenthal CK, et al., 2009, Scheffler IE, 2008, Titford M, et al., 2005) 미토콘드리아연구의역사 현미경과조직염색기술덕택으로미토콘드리아가명명되고존재가증명되었다면, 형광현미경과생체염료덕택으로미토콘드리아의상세구조가밝혀졌다. 형광염료가붙은항체를이용한염색처리를하면형광현미경을통하여미토콘드리아를관찰할수있었다. 또한로다민 2
14 유도체나 다른 친지질성 생체염료를 적용하면 살아있는 세포에서 미토콘드리아의 생활사를 관찰할 수 있었다. 그 원리로는 미토콘드리아막의전위차를이용하여물질을내강으로운반되는기전을 이용하여 미토콘드리아 안으로 염료를 농축시키는 것이다. 이를 저속으로촬영하면미토콘드리아의역동적인활동 ( 크기와모양의변화 ), 세포질 안에서 미토콘드리아의 분포, 세포골격을 동요시켜 미토콘드리아를 재배치 등을 파악할 수 있었다. 이러한 기법을 통해 미토콘드리아의 특성들이 밝혀졌다. 간세포나 섬유모세포에서 관찰된 미토콘드리아는소시지모양으로길이 3-4 μm, 지름 1 μm미만으로 모양이나크기는작은세균과유사한크기였다. 미토콘드리아의수는 세포의종류에따라수백개에서수천개에달할만큼달라질수있다. 예를 들면 인체의 난자는 100,000개 이상의 미토콘드리아가 있고, 정자에는 거의 없다. 또한 미토콘드리아의 성장과 분열은 각 세포의 분화와기능에따라다르다는점이발견되었다. (Scheffler IE, 2008) 이러한미토콘드리아의구조에대한연구뿐만아니라미토콘드리아의 기능에대한연구도 `1900년대전반부에이루어졌다. 미토콘드리아는 호흡과관련이있고세포내에너지를생산하는역할을담당하고있음이 밝혀졌다. 근육조직에대한연구를통해미토콘드리아안의시토크롬, 철결합물, 플라빈과 피리딘 뉴클레오티드, 크렙스 회로 등에 대한 정보가발견되었다. 이외에도조직을갈아서얻어진미토콘드리아에서 직접적으로에너지생산을관장하는효소들을정제하여연구해보려는 시도가 있었으나 세포들을 깨뜨린 부유액에 대한 완충제와 배지가 부적합하였기때문에좋은성과는얻지못했다. 1940년대에이르러서야 원심분리기를 이용하여 세포내 구조물을 분획할 수 있는 분석기법이 개발되어, 이를 통해 생화학적인 특성에 대하여 분석하고 연구할 수 있었다. 간세포를이용한연구에서지방산산화효소가미토콘드리아와 관련이있음이밝혀졌다. (Andrews RM, et al., 1999) 이러한미토콘드리아구조와기능에대한연구가진행되고있을때에 1963년 미토콘드리아의 연구 역사에 또 하나의 획기적인 발견이 이루어졌다. 미토콘드리아 안에는 세포핵 디옥시리보핵산 (DNA) 와 별도로독립적인 DNA가존재함이이때발견된것이다. 이미효모균 연구를통해멘델의유전법칙을따르지않는세포질유전이있음이 밝혀지면서, 미토콘드리아만의 독립적인 유전물질이 있을 것이라는 예측은 있었고, 마침내 그 존재를 증명할 수 있게 되었던 것이다. 인체에서는 1967년에서야 비로소 처음 그 구조가 밝혀졌다. 이 3
15 발견으로세포핵유전자와미토콘드리아의유전자가세포안에서어떻게작용하는지에대해서새롭게이해하게된계기가되었다. 또한이발견은미토콘드리아의유래가공생에서기원하였다는가설을뒷받침해주는증거로서도의미가있었다. (Andrews RM, et al., 1999, Clayton DA, 1992, Moraes CT, et al., 2002, Scheffler IE, 2008) 미토콘드리아 DNA 염기서열에는다양한변이가발생할수있고, 이렇게시간이흘러감에따라변이가축적될것이며, 이렇게축적된변이는분자생물학적인시간의척도가될수있을것으로보아미토콘드리아 DNA 염기서열비교를통해인류의기원과이동에대해추정해보는연구가시작되었다. 이러한연구를통해밝혀진유전적다양성은유전학, 계통발생학등에서의연구주제였을뿐만아니라법의학개인식별영역에서도관심을갖기시작하였고, 유전적다양성을개체식별에응용하였다. 1981년에인체의미토콘드리아 DNA의표준전체염기서열이발표되었으며, 이를 Cambridge Reference Sequence, CRS 로명명하였다. 이염기서열은인류뿐만아니라모든생물종에서최초로인류의미토콘드리아 DNA 염기서열을밝혀낸연구이기도하였다. 다만그때당시모든유전자부위가다해독된것은아니었고, 남아있던밝혀지지않았던유전자는 1986년에이르러서야그해독이완성되었다. 1999년에는 CRS와다른연구결과에서알려진미토콘드리아 DNA 염기서열과비교해보았을때, 일부뉴클레오티드에차이가있음이밝혀졌다. 이러한차이는실험적오류및드문다형태 (polymorphism) 로보고, 이를개정하여 revised Cambridge Reference Sequence, rcrs 라고명명된개정표준미토콘드리아 DNA 염기서열을발표하였다. 2012년에는개정캠프리지표준서열이유럽인의하플로그룹에속한것으로알려지면서계통발생학적특성을담아개정한개정된인류표준서열 (Reconstructed Sapiens Reference Sequence) 가제안되었으나, 여러연구에서 rcrs를기준으로분석해왔기때문에아직도 rcrs를표준서열로사용되고있다. 이러한표준염기서열및유전자의해독정보는법의학의개체식별뿐만아니라하플로그룹 / 하플로타입연구에따른계통발생학연구, 미토콘드리아유전학적연구, 질병학적연구의중요한계기가되었다. (Behar DM, et al., 2012, Alverts B, et al., 2008, Anderson S, et al., 1981, Andrews RM, et al., 1999, Bandelt HJ, et al., 2006, Chomyn A, et al., 1986, Scheffler IE, 2008, van Oven M, et al., 2009,) 4
16 1.2. 미토콘드리아유전시스템 미토콘드리아의성장과분열과미토콘드리아 DNA 미토콘드리아는세포내구조물이지만세균같은미생물처럼기존에존재하던미토콘드리아에서새로운미토콘드리아로성장하거나새로이분열되는방식으로태어난다. 세포분열이일어나는세포안에서는미토콘드리아의수는두배가되었다가각딸세포에각각동등하게분배된다. 세포분열을하지않는세포일지라도미토콘드리아의수명이다하거나좀더많은수의미토콘드리아를필요로할때미토콘드리아의성장과분열을통해이러한수요를충족시킨다. 이에대해서는실제살아있는세포에서미토콘드리아를관찰해보았을때역동적이어서자주분열하고융합하며모양을변형시킴이밝혀지면서증명되었다. 미토콘드리아의수와모양은여러가지다양한생리적조건에따라시시각각달라지며심지어동일세포안에서조차도미토콘드리아의분열과융합의비율에따라상대적으로변화한다. 이러한미토콘드리아의성장과분열은두개의축으로구성된유전시스템이관여하는복잡한과정을거친다. 그시스템의한축은미토콘드리아안에세포핵 DNA와는독립적으로존재하는 DNA, 이에따라리보핵산 (RNA) 를합성하고단백질합성을하는자체유전시스템이고, 다른한축은세포핵 DNA의유전시스템이다. 즉미토콘드리아유전체의유전시스템을통해전사및합성된단백질과세포핵유전체의유전시스템에서전사및합성된단백질이상호작용하여기능함으로서미토콘드리아성장과분열에관여한다. 세포핵 DNA를통해합성된단백질은미토콘드리아내로운반되지만, 미토콘드리아내에서합성된단백질은세포질로나가지않고그대로머무르기때문에이들단백질은일방통행의움직임을보인다. 예외적으로세포질로단백질을내보낼때가있지만이는오직세포자멸사가진행되는경우에서만일어난다. (Hayashida K, et al., 2005, Scheffler IE, 2008) 미토콘드리아 DNA 복제 미토콘드리아의성장과분열에중요한역할을하는미토콘드리아의 5
17 유전시스템에있어서미토콘드리아 DNA의복제는세포핵 DNA가복제되는시기인세포분열주기의 S기에서만국한되어일어나지않고, 세포분열주기와상관없이일어난다. 미토콘드리아안에는여러복제수의 DNA가있지만이들중에서복제하려는 DNA는무작위로선택된다. 따라서어떤미토콘드리아 DNA는세포분열주기동안한번이상여러차례복제되는반면어떤미토콘드리아 DNA는전혀복제가일어나지않는다. 그럼에도불구하고매세포분열주기마다미토콘드리아의 DNA의전체양은 2배가되도록일정하게조절되고있으며, 또한각딸세포에동일한양의미토콘드리아 DNA가분배되도록조절된다. 미토콘드리아 DNA가복제되는장소는세포핵근처에있는미토콘드리아의내부이다. 핵을제거한세포에서는미토콘드리아의복제가완전히정지되는것을관찰한실험을통해증명되었으며, 이를통해미토콘드리아 DNA 복제에필요한단백질이세포핵 DNA에도암호화되어있음을발견하게된계기가되기도하였다. (Alberts B, et al., 2008, Bogenhagen D, 1977, Bogenhagen DF, 2012, Scheffler IE, 2008) 미토콘드리아 DNA 변화 미토콘드리아 DNA의복제가일어날때, 이상적으로는염기서열정보가그대로복제되어야할것이다. 그러나실제로는복제과정에서미토콘드리아 DNA 염기서열에돌연변이가발생하거나 DNA 복제수에변화가발생한다. 이러한 DNA의변화를유발하는원인으로는외적원인과내적원인이있다. 외적원인에는주로미토콘드리아단백질과막의지질성분의산화인산화가일어나면서수반하는부산물 ( 활성화산소등 ) 이있다. 미토콘드리아 DNA는지정학적으로산소를이용하는에너지대사가활발하게일어나는미토콘드리아내부에함께위치하고있으며이대사과정에서활성화산소가생성되는곳과근접하여위치하고있다. 또한히스톤과같은보호기능을해주는단백질의부재로미토콘드리아 DNA는산화에의한손상에매우취약할수밖에없다. 특히인체의경우, 활성화산소에의해유발된미토콘드리아 DNA 염기서열의손상은세포핵 DNA의손상에비해훨씬더광범위하면서오래유지된다. 활성화산소는염기뿐만아니라 DNA의뼈대에해당하는당인산염에도손상을줄수있고, DNA 가닥이끊어지거나가교 (crosslink) 를형성하기도한다. 그외외인성원인으로여러가지 6
18 유전자독성물질 ( 산업물질, 자외선, 방사선, 흡연, 화학물질, 환경독성물질, 약물등 ) 이있다. 미토콘드리아내막은음전하를띄고, 많은독성화학물질들은양전하를띄는친지질성물질이기때문에, 이물질들이미토콘드리아내부로축적되고, DNA는손상받기쉬운상태가된다. 내적원인으로는미토콘드리아 DNA가복제되는과정에서자연적으로발생할수있는오류로인하여생성된점돌연변이나결실등이있다. 미토콘드리아에만있는 DNA 중합효소 γ는미토콘드리아 DNA의복제와복구를담당하는데, 내적원인에의한손상은이효소의착오로발생하게된다. (Akhmedov AT, et al., 2015, Alberts B, et al., 2008) 미토콘드리아 DNA 복구시스템 연구초기에는미토콘드리아 DNA염기서열의변화가발생할때그에대한복구시스템은없을것으로여겨졌다. 그이유는손상받은미토콘드리아 DNA 전체를분해를시키고손상받지않은 DNA를주형으로새로 DNA를복제하여기존의손상받은 DNA를대신하기때문이다. 각세포당수백개에서수천개복제수의손상받은 DNA가분해된다. 그러나이후수년에걸친연구를통해미토콘드리아에도세포핵 DNA 복구시스템과거의유사한 DNA 복구시스템이있음이밝혀졌다. 그종류로는 염기절단복구 (base excision repair), DNA 가닥복구, 불일치염기복구, 상동재조합 (homologous recombination), 비상동말단봉합 (nonhomologous end joining) 등이있다. 다만미토콘드리아에는세포핵 DNA 복구시스템과달리뉴클레오티드절단복구시스템이없다. 복구시스템에관여하는단백질은모두세포핵 DNA에암호화되어있어서세포핵에서전사및합성되어미토콘드리아내로운반된다. 염기절단복구에는짧은서열복구과긴서열복구등두종류가있다. (Akhmedov AT, et al., 2015, Alberts B, et al., 2008) 미토콘드리아 DNA 변화와복구시스템의불균형및 유전시스템에미치는영향 DNA 염기서열의복제수의변화나돌연변이를방지하기위해분해및 복제, 또는복구시스템이존재함에도불구하고, 미토콘드리아 DNA 는 7
19 세포핵 DNA보다 10배이상으로많은뉴클레오티드치환이발견된다. 그이유는첫째미토콘드리아의 DNA 복제과정에서정확성이떨어지고, 둘째 DNA 복구는비효율적이기때문이다. 미토콘드리아 DNA 전체염기서열의복제와복구, 미토콘드리아유전체의전사, 단백질합성과정에서오차율은상대적으로높고정확성이떨어진다. 그럼에도불구하고미토콘드리아유전시스템에서형성된생산물은상대적으로거의손상되지않는데, 이는세포핵유전시스템에비해미토콘드리아유전시스템이상대적으로단순하기때문이다. (Akhmedov AT, et al., 2015, Alberts B, et al., 2008) 미토콘드리아가이런별도의유전시스템을유지하기위해서는리보솜단백질, 합성효소들, 진행및수정에관여하는효소등이시스템에관여하는 90여개이상의단백질합성이필요하고, 이를위해핵유전체에별도로동일한수의유전자를통해암호화시켜야한다는점에서비효율적으로보인다. 이에대해서초기원시세포와포식당한세균의공생의개념을적용해본다면, 미토콘드리아의 DNA가오랫동안점진적으로세포핵 DNA로전달되어왔고이제는전달과정의마무리단계에있을것으로보인다. 만약세포핵 DNA로의전이를완전히마치면미토콘드리아의유전시스템은기능을상실할것으로생각해볼수있을것이다. (Akhmedov AT, et al., 2015, Alberts B, et al., 2008) 1.3. 미토콘드리아 DNA 의유전학적특성 미토콘드리아 DNA 의유전방식 : 모계유전 미토콘드리아 DNA의유전은세포핵 DNA의유전법칙인멘델의유전법칙을따르지않는다는특징은효모균연구를통해알려진비멘델유전또는세포질유전으로서알려져있었다. 다만효모군의미토콘드리아의유전과인체의미토콘드리아다른점이라면효모균의경우부모양쪽에서반반씩미토콘드리아 DNA를받는다는점일것이다. 인체의경우미토콘드리아 DNA를모계유전으로보는이유는다음과같다. 첫째난자와정자가수정할때, 난자세포만이세포질을구성하고정자는거의기여하지않아서모계한쪽에서만미토콘드리아 DNA를 8
20 전달하게된다. 수정된난자는약 2,000여개의복제된미토콘드리아를가지고있으며, 한두개를제외한나머지모든미토콘드리아는모두모계에서유전된것이다. 다른연구에의하면성숙한난자는수천개의미토콘드리아가있고, 미토콘드리아 DNA는약 100,000여개이상의복제수가있다고하였다. 둘째는특이적으로부계에서오는미토콘드리아를인지하는기전이있어서난자와정자가수정될때들어오는소수의부계미토콘드리아를제거하기때문으로본다. 수정란안에있는부계미토콘드리아는 4세포분할기와 8세포분할기를거치면서유비퀴틴에의해수정된다. 유비퀴틴신호는남성생식기의정자미토콘드리아에서도확인되는점으로비추어수정전에이미정자의미토콘드리아는선택적퇴행을위해유비퀴틴으로꼬리표를붙인것 으로설명하였다. 수정후에유비퀴틴으로표시된부계미토콘드리아는단백질분해효소복합체 (proteasome) 나용해소체에의해분해되는것으로보인다. 미토콘드리아의모성유전은동물에서먼저관찰되었고, 인간에서는 1980년처음 3가족을대상으로제한효소를이용한기법으로미토콘드리아 DNA를비교분석하였다. 가계도를분석하였을때, 멘델의유전법칙즉우성또는열성유전과는다르고반성유전과도맞지않았고, 모계를통해나타나는모성유전임을증명하였다. (Alberts B, et al., 2008, Giles RE, et al., 1980, Hayashida K, et al., 2005, Holland MM, et al., 1999, Sato M, et al., 2013, Scheffler IE, 2008) 드물지만인체에서부성미토콘드리아 DNA가있다는문헌보고가있었고인체에서도양측부모의미토콘드리아유전체의제조합은과연일어나지않는것인지에대한의문이없었던것은아니다. 그러나현재까지는여러연구및연구자들사이에서미토콘드리아의클론형성능 (clonality) 증개체내에서미토콘드리아 DNA는동형세포질성이라고간주한다. 또한인류에서미토콘드리아 DNA 재조합이일어난다고고려할만한충분한과학적근거는아직알려진바가없기도하였다. 1997년의연구에서법의학실무에서사용하는같은방법 ( 증폭시킨 DNA를직접염기서열분석하는기법 ) 을적용하여 69쌍의아버지와아들의미토콘드리아 DNA 염기서열을직접비교분석해보았을때부계미토콘드리아 DNA가있었던사례는없었다. 만약이형세포질성 (heteroplasmy) 이부계에서온미토콘드리아 DNA라면, 전혀친족관계가없는무작위로선택된 2명의코카시안인종을대상으로미토콘드리아 DNA의염기서열을비교분석하였을때, 9
21 일반적으로조절부위에서 8개이상의변이가관찰되는점을감안하였을때, 부계에서전달되어혼합된 DNA에서는더많은부위에서변이가관찰되어야할것이다. (Budowle B, et al., 2003, Coble MD, et al., 2004, Elson JL, et al., 2006, Hayashida K, et al., 2005, Holland MM, et al., 1999, Parsons TJ, et al., 1997, Scheffler IE, 2008, Schwart M, et al., 2002) 미토콘드리아 DNA 유전과이형세포질성 앞서설명한대로미토콘드리아 DNA 복제과정에서돌연변이등변화가발생하고, 세포핵 DNA보다오차율이훨씬더높다. 이러한특성덕택으로, 각개체에서각기다른고유의다양한 DNA로나타날수있고각각의변이는독립적으로복제되고분리될수있으며, 모계를통해후손에게이러한변이를전달할것이다. 특히돌연변이의여러유형중대단위의결실은유전이되지않지만점돌연변이는대개유전된다. 따라서각개체의미토콘드리아 DNA에는각모계조상에서물려받은각기고유한 유형 이있을것이다. 그러나이러한모계의고유한유형과구별되는또다른고유한유형이있음이밝혀졌다. 미토콘드리아 DNA 세대간에전달하는과정에서변이가전달되는정도를제한하는 유전적병목 (genetic bottleneck) 기전이있음에도불구하고, 동일개체안에서 2개이상의미토콘드리아 DNA 유형이혼합되어있는현상, 즉이형세포질성 (heteroplasmy) 이발견되었다. 다시말하면각개체를구성하는각세포안의미토콘드리아 DNA에서살아가는동안돌연변이가제각각발생할것이다. 그리고각자돌연변이가발생한미토콘드리아 DNA와돌연변이발생이없는미토콘드리아 DNA의비율은각세포마다다르게나타날뿐만아니라동일개체내의각장기조직마다도다르게나타날것이다. 이형세포질성에대한초기연구는망막을가지고진행하였는데, 망막이신경조직의연장으로서노화에따라미토콘드리아 DNA의돌연변이가증가하는점에착안하여연구를하였으나이행세포질성에있어서는개체내변이는아주적으며, 따라서각개체는동형세포질성 (homoplasmy) 으로간주할수있을것으로보았다. 이후여러연구들에서이러한관점은이후상당기간계속지속되었는데아마도당시염기서열분석기법의수준으로는이형세포질성을배경잡음과구별하기어려웠기때문이기도했을것이다. 그러나과학기술의발달로 10
22 개선되거나새로운염기서열분석기법의도입으로이형세포질성을더잘 발견할수있었다. 다만현저하게나타난뉴클레오티드만변이검출로 인정하고 나머지는 원인 미상의 불확실한 염기로 간주하였었다. 니콜라스 II세황제의유골에대한법의학감정이진행되던중발견된 점이형세포질성 (point heteroplasmy) 을기점으로이형세포질성에대한 연구가이어졌다. 일반적으로이형세포질성의빈도는비교적으로낮아서 인구의 2-8% 정도로알려졌다. (Alberts B, et al., 2008, Bandelt HJ et al., 2006, Bendall KE, et al., 1996, Hasegawa M, et al., 1993, Hauswirth WW, et al., 1982, Holland MM, et al., 1999, Koehler CM, et al., 1991, Parsons TJ, et al., 1998, Poulton J, et al., 2000, Scheffler IE, 2008, Wakeley J, 1993) 이어진여러연구를통해이형세포질성의형성과각수준이변화하는 기전이 발견되었다. 미토콘드리아 DNA의 복제와 분해는 세포분열에 따른 세포핵 DNA의 복제와 상관없이 계속적으로 일어나고 각각의 복제나분해의대상은무작위로선택된다고앞서설명하였다. 이러한 특징은세포분열이거의없는것으로알려진근육세포나뇌신경세포에서 왜 이형세포질성의 수준이 다르게 나타나는지, 그리고 미토콘드리아 DNA의 돌연변이에 따른 질병의 특성을 이해하게 해준다. 또한 영양분리 즉 세포질의 무작위 또는 불공평한 구획과 분리 는 혈액세포와 같은 계속 증식하는 세포에서도 이형세포질성 수준이 달라지게하였다. 뿐만아니라 유전적병목 을통해이형세포질성이 후손에게전달됨으로서모계의유형과는또다른유형이나타날수있게 되었다. 따라서 이형세포질성은 모계에 해당하는 친족사이에서의 차이뿐만아니라동일개체내의각장기및조직의세포사이에서도 다른이형세포질성비율이나타날수있다는것이다. 병목 이론은 암소에 대한 연구를 통해 제시되었다. 동형세포질성에서 이형세포질성으로, 동형세포질성에서 다른 동형세포질성으로의 변화가 암소의 각 세대에서 발생한다는 것이 발견되었다. 이러한 현상을 설명하기 위하여 난자의 발생 과정 중 어떤 단계, 또는 생식세포 발달과정동안미토콘드리아 DNA의수가비교적적은수로줄어드는 병목 이론이 제시되었다. 즉, 병목현상 초기에 일어나는 현상처럼, 미토콘드리아 DNA 일부 군이 전달될 것이다. 그리고 전달된 미토콘드리아 DNA는성숙난자에서최대 100,000 가량의복제수로 복제될것이고이과정에서원래의미토콘드리아 DNA 염기서열과다른 분율의 이형세포질성 변이가 나타날 것이며, 이후 진행되는 11
23 감수분열과정중에또다른분율의이형세포질성변이를지닌채분리된다. 인체의난소발생과정에서도이러한미토콘드리아 DNA 병목현상이발생함이관찰되었다. 쌍둥이와그가계를대상으로연구하였을때세대사이에다른분율의이형세포질성변이를보임이관찰되었다. 감수분열과정중병목을거치면서 2에서 100개사이의미토콘드리아 DNA 복제수 ( 평균 3-20) 가전달되는것으로추정하지만, 실제병목을통해전달되는미토콘드리아 DNA의양은일정하게전달된다고하기보다는매전달단계에서확률적으로달라질수있다. 이렇게유전을통해전달받은미토콘드리아 DNA의이형세포질성은각개체내에서장기마다그수준이무작위로다르게나타날수있게된다. 결국이형세포질성의수준은미토콘드리아 DNA를오랜시간보존및유지하는관련인자들을통하여각개체의미토콘드리아유전체수준에서어느정도조절될것이다. 이뿐만아니라세포핵의유전자와관련된인자도이형세포질성의수준의조절에관여한다. 이러한특징은미토콘드리아 DNA 돌연변이와관련된질병의발병기전을이해할수있게하며, 미토콘드리아 DNA 돌연변이의유전적전달과인류의진화를이해하는데에도중요할것이다. (Alberts B, et al., 2008, Bandelt HJ et al., 2006, Bendall KE, et al., 1996, Hasegawa M, et al., 1993, Hauswirth WW, et al., 1982, Holland MM, et al., 1999, Koehler CM, et al., 1991, Parsons TJ, et al., 1998, Scheffler IE, 2008, Wakeley J, 1993) 이형세포질성에는하나의염기부위에다른염기가치환되어형성되는점이형세포질성뿐만아니라수개의염기부위에연속적으로변이가형성된길이이형세포질성 (length heteroplasmy) 이있다. 길이이형세포세포질성은과변이부위의다시토신뻗기부위 (polycytosine stretches) 에서흔하게나타난다. 과변이부위 I의 16189번위치의뉴클레오티드는티민 (T) 이어서각양쪽의시토신 (C) 의흐름을끊어주는역할을한다. 그러나이부위가 T에서 C로변이가발생하면 C가연속적으로배열되기때문에정확성이낮은것이특징인미토콘드리아 DNA 중합효소에의해 C의연속적인배열이생성된다. 이는각개체마다다른길이의 C 뻗기가형성됨으로써염기서열분석에서오차가나는부분이기도하다. 길이이형세포질성은과변이부위 II의 C뻗기에서도나타날수있지만, 이경우쉽게알아볼수있는뚜렷한길이변이가있다. (Alberts B, et al., 2008, Bendall KE, et al., 1996, Hasegawa M, et al., 1993, Hauswirth WW, et al., 1982, Holland MM, 12
24 et al., 1999, Koehler CM, et al., 1991, Parsons TJ, et al., 1998, Scheffler IE, 2008, Wakeley J, 1993) 이형세포질성에대한연구를통해발견된중요한점은미토콘드리아 DNA의각뉴클레오티드위치는동일한확률로이형세포질성이발생하는것이아니라, 자주호발하는특정부위 ( hot spot ) 가있다는점이다. 이형세포질성이자주발생하는염기서열부위는인류가생존해온오랜시간동안변이가형성되고축적되어결국각개체사이의차이를만드는염기서열부위에상관관계가있을것이었다. 러시아의황제니콜라스 II세의유골의법의학감정사례에서보여주듯이이형세포성질이미토콘드리아 DNA 분석을이용한개인식별에있어서식별력을증강시키는데유용할것으로생각된다. 다만이경우개체의발달과생존기간동안개체내에서발생하는변이와유전적으로전달되는변이와구별되어야할것이다. 일부사례에서는드물기는하지만, 원래의뉴클레오티드가대부분변이로치환되어동형세포질성으로보일만큼큰차이를보이기도해서개인식별감정에서개체식별에대한해석및감정에있어서어려운문제가될수있다. 그외이형세포질성의수준이질병의발현에영향을줄수있는점도임상의학적으로중요한부분이다. (Alberts B, et al., 2008, Bendall KE, et al., 1996, Hasegawa M, et al., 1993, Hauswirth WW, et al., 1982, Holland MM, et al., 1999, Koehler CM, et al., 1991, Parsons TJ, et al., 1998, Scheffler IE, 2008, Wakeley J, 1993) 1.4. 미토콘드리아 DNA 의구조 미토콘드리아내 DNA 구조 미토콘드리아의 DNA는원형의이중나선구조로되어있으며, 질병등비정상적인상태에서는이합체 (dimer) 로형태가바뀌기도한다. 또한미토콘드리아 DNA는미토콘드리아구조의영향을받는다. 미토콘드리아의내부공간에는여러작은공간들로나뉘어있는데, 그러한공간들을나누는격막은내막으로형성되어있다. 이들각공간에는여러복제수의미토콘드리아 DNA가들어있으며, 이를핵모양 (nucleoid) 라고하는데, 세포핵내염색질보다는세균의 DNA와 13
25 유사하여히스톤이없다. 미토콘드리아핵모양은비교적안정된유전물질로인체세포에서는하나의핵모양당평균 1.4개 7.5개의미토콘드리아 DNA로복제수로구성되어있으며이들 DNA를코팅하는것처럼결합하는단백질이있다. 따라서미토콘드리아내부에이러한작은공간들의형성의정도는가지고있는미토콘드리아 DNA의복제수에따라달라진다. 미토콘드리아 DNA의복제수가많다는것은일부 DNA에돌연변이가생겼을때그로인한유전자기능의공백을다른정상 DNA가상보적으로대신기능해주는등의보호역할을담당함을의미한다. 미토콘드리아핵모양과관련된단백질에는미토콘드리아 DNA와결합하는단백질 (single strand DNA binding protein), DNA 나선구조의나선을조절하여핵모양내에서미토콘드리아 DNA의압축정도에관여하는단백질 (high mobility group box protein TFAM), 핵모양의안정성유지에관여하는 prohibitin 단백질, TFAM 단백질을분해시켜서미토콘드리아 DNA 복제수를조절하는데관여하는 Lon 단백질분해효소, 리보솜관련단백질등이있다. 또한세포골격을유지하는단백질이미토콘드리아기질내로들어와미토콘드리아핵모양과접촉하는것이알려졌는데아직그의의에대해서는아직밝혀지지않았다. (Alberts B, et al., 2008, Bogenhagen DF, 2012, Scheffler IE, 2008) 미토콘드리아의 DNA 가닥의구조와구성 인체에서미토콘드리아의 DNA는 16,569개의뉴클레오티드쌍으로 구성된원형의 DNA로 13개의단백질을합성하는유전자가포함되어 있으며, 2개의리보솜RNA 구성성분과 22 전이RNA들이있다. 주요 염기구성에서구아닌과시토신등의염기가비대칭적으로분포하는점 때문에 무거운 가닥 (heavy chain) 과 가벼운 가닥 (light chain) 으로 구분한다. 바깥쪽 DNA가닥이 무거운 가닥이고, 안쪽 DNA가닥이 가벼운 가닥에 해당하고, 바깥쪽 DNA에 주로 유전자가 암호화 되어있다. ( 그림 1 참조 ) 이러한 16,569개의뉴클레오티드로구성된 DNA는 세포핵 DNA에 비해 비교적 작은 크기의 유전체이다 보니 초기에는 DNA 염기서열분석 연구 대상으로 최적이었다. 1981년 미토콘드리아 DNA를구성하는전체뉴클레오티드를완전히분석하여 발표하였다. 당시 알려진 미토콘드리아 전이RNA 염기서열과 부분적으로 알려진 아미노산 서열과 함께 비교 분석함으로서 원형의 14
26 DNA 지도가완성되었다. 인체의미토콘드리아 DNA를세포핵 DNA, 다른유사한엽록체의 DNA와세균의 DNA 등과비교해보면, 거의모든뉴클레오티드는단백질이나전이RNA를암호화하고있는염기서열의일부로여겨지며, 이러한암호부위는서로밀접하여연이어있어서조절부위염기서열은적은비율로구성되어있다. 또한미토콘드리아단백질을합성하기위해서는겨우 22개의전이RNA가필요해서코돈과대응코돈이짝을이루는부분이느슨하게되어있어서많은전이RNA가세번째동요부위 (Wobble position) 의뉴클레오티드들중하나를쉽게인지하게하여전이RNA가결국결합하게되고, 이러한짝형성은적은양의전이RNA만으로도단백질합성이일어날수있게한다. 마지막으로유전자암호의의미가다양하다는것인데미토콘드리아유전자염기서열과그에대응하는단백질의아미노산서열을비교분석해보면, 64 코돈중 4개의코돈이같은코돈임에도다른아미노산으로전사된다. (Alberts B, et al., 1008, Anderson S, et al., 1981, Chomyn A, et al., 1986, Clayton DA, 1992, Garrigan D, et al., 2006, Moraes CT, et al., 2002, Scheffler IE, 2008, Wills C, 1995) 미토콘드리아의 DNA는유전자정보가있는암호부위 (coding region) 와유전자가아닌염기서열이모인부분인조절부위 (control region) 로구성되어있다. 조절부위는길이약 1,125 염기쌍으로구성되어있으며, DNA의복제가시작되는부위일뿐만아니라, RNA 전사의시작부위 (promotor) 가있는부위이다. 1981년에발표된캠브리지표준염기서열 (Cambridge reference sequence, CRS) 은인위적으로조절부위중앙에서부터염기서열번호를시작하였기때문에, 조절부위는 1번에서 576번사이와 16,024번에서 16569번사이에위치한염기서열을말하며, 나머지염기서열 (577번에서 16023번까지 ) 이암호부위에해당한다. (Anderson S, et al., 1981) 암호부위의유전자의순서는안정적으로유지되어왔으나개별유전자사이의뉴클레오티드의염기서열은종마다현저하게다르며특히인류에서는상당한염기서열변이의다양성즉다형태 (polymorphisms) 가있음이관찰되었다. 이러한점때문에인류학뿐만아니라법의학에서이부위에대해많은관심을기울이고있다. (Holland MM, et al., 1999, Scheffler IE, 2008) 세포핵의미토콘드리아 DNA 염기서열 (numt) 15
27 미토콘드리아 DNA는미토콘드리아내에서만생장하는것이아니라세포핵 DNA의염색체내에서여러조각으로있으면서생장한다. 이는인체뿐만아니라미생물, 동물, 식물등여러연구에서확인된현상으로, 세포핵의 DNA 염기서열중에서미토콘드리아 DNA와거의유사한부분이있어서이를세포핵 DNA에삽입된미토콘드리아 DNA 즉세포핵의미토콘드리아 DNA (numt; nuclear inserts of mitochondrial DNA) 라고불리운다. 이는인류의진화및이주를이해하고, 고대시료의미토콘드리아 DNA를해석하는데있어서미토콘드리아 DNA와 numt를구별하는것이중요하지만또한질병의발생기전을이해하는데에도중요한부분이었다. 인체에서는각염색체뿐만아니라전체염색체에서 612개의 numt가다양한염기서열의길이로서분포하고있고, numt는적어도전체세포핵 DNA의약 0.016% 를차지할것으로추정하였다. 세포핵 DNA내로삽입되는기전으로는미토콘드리아 DNA 그자체의삽입, RNA 중간매개체에의한삽입에의해발생할것으로본다. (Bandelt HJ et al., 2006, lopez JV et al., 1994, Woischnik M et al, 2002) 2. 법의학에서미토콘드리아 DNA 의의의 2.1. 법의학에서활용되는미토콘드리아 DNA 의특성 미토콘드리아 DNA 와세포핵 DNA DNA에대한과학적지식이축적되고분석기법이발달함에따라이를 사법절차에도입하여활용하기시작하였다. 생물학적증거물로서세포핵 DNA는많은정보를줄수있다는점에서최적의대상일것이다. 다만 세포핵 DNA와 비교하였을 때 미토콘드리아 DNA가 지닌 몇가지 장점이있어서여전히유용한정보를얻을수있는부분이있다. 첫째, 하나의 세포에서 세포핵 DNA는 두 개의 복제수 밖에 없지만, 미토콘드리아 DNA는평균적으로수천개의복제수를가지고있어서, 훨씬 더 민감한 분석이 가능하다는 점이다. 둘째로 미토콘드리아 DNA는 양쪽 부모에서 물려받지 않고, 재조합이 일어나지 않는 16
28 모계유전이라는점이다 법의학실무에서미토콘드리아 DNA 분석의장점 변질이진행된시료나오래된유골에서처럼세포핵 DNA가변성되는경우, 손톱, 머리카락처럼세포질만남아있고세포핵이없어서아예분석이불가능한경우에서처럼세포핵 DNA를확보할수없는경우에서도세포질은남아있기때문에미토콘드리아 DNA는확보할수있다. 따라서앞서설명한미토콘드리아 DNA의장점들을적용하여추가적인정보를확보할수있으며, 또한모계의대조시료가있는경우확보된미토콘드리아 DNA 정보를이용하여개인식별, 법생물학증거물에대하여추가적인해석이가능해진다. (Coble MD, et al., 2004, Coble MD, et al, 2009, Giles RE, et al., 1980, Scheffler IE, 2008, Seo Y, et al., 2000) 개체식별측면에서법의학에서미토콘드리아 DNA 의특징 미토콘드리아 DNA의변화는각개체마다다르게나타나는다형성을만들고이는미토콘드리아 DNA 염기서열중특정부위에서높게나타난다. 암호부위에비해조절부위에서각개체사이의다형성이높은비율로발생하는데, 이는암호부위에비해조절부위염기서열에서선택적제약이상대적으로낮기때문으로추정된다. 특히이러한조절부위의다형성이높게나타나는부분을과변이부위라고하며, 과변이부위 I은 16024번뉴클레오티드와 16365번뉴클레오티드사이에위치하고있으며, 과변이부위 II는 73번뉴클레오티드에서 340번뉴클레오티드사이에위치하는데이부위들의위치는고정되어있지는않고연구에따라조금씩달라지기도한다. (57번에서 372번사이 ) 이러한염기서열의변이가진화의과정에서형성된미토콘드리아 DNA의유전적다양성으로정의되며이러한유전적다양성을보이는수많은염기서열을모은데이터베이스및하플로그룹 (haplogroup) 분류의개념이정립되었다. 또한이러한각개체사이의염기서열변이차이의특성이개인식별의근간이되었으며실무적으로조절부위를중심으로염기서열분석을시행하게된근거가되기도하였다. (Holland MM, et al., 1999, van Oven M, et al., 2009) 17
29 2.2. 미토콘드리아 DNA, 계통발생학, 하플로그룹 (haplogroup) 미토콘드리아 DNA 와인류의진화연구 미토콘드리아 DNA를인류의진화연구에적용한것은 1987년에발표된연구였다. 이연구에서여러대륙에서유래한인종 (34명의아시아인, 21명의호주원주민, 26명의뉴기니아원주민, 46명의코카서스인, 20명의아프리카인 ) 으로구성된 147명의사람들을대상으로하여미토콘드리아 DNA에서제한효소가작용하는부위를분석하여 DNA 지도를완성하였다. 이미다른선행된다른연구들을통해분자유전학기법을이용한미토콘드리아 DNA 염기서열분석을통해인류의기원과진화에대한연구가가능할것이라는예상이없었던것은아니었다. 그러나여러다양한인구를대상으로분석한이연구결과를통하여미토콘드리아 DNA 분석을통한접근방법이현생인류와진화와기원, 우리조상들의이주형태에관한의문에대하여과학적으로접근가능함을보여주었던획기적인계기가되었다. 이모든미토콘드리아 DNA는약 200,000년전에아마도아프리카에서살았을것으로추정되는한여성에서유래되었다. 라고하는이러한기술은이연구논문의초록에기술되었던것으로, 이를여러매체에서문자그대로인용하여논란을불러일으켰을만큼널리알려진인용구가되기도하였다. (Brown WM, et al., 1982, Cann RL, et al, 1987, Scheffler IE, 2008) 이후분자생물학적기술의발전으로더많은종류의제한효소를사용할수있게되었고, 중합효소연쇄반응기법의적용, 특정부위의직접적인염기서열분석기법적용등에의해더세밀한분석이가능하게되었을뿐만아니라, 지구상의가장먼지역의인류미토콘드리아 DNA가분석되어인류의기원을설명하고자하는노력들이이어졌다. 미토콘드리아 DNA의진화의비율은세포핵 DNA의진화비율에비해더큰규모로발생할것이다. 따라서미토콘드리아 DNA가인류의진화를이해하는데적합한상세한시간척도가될수있다. 미토콘드리아 DNA의염기서열변화는시간이흘러감에따라축적되고, 같은인종군에서다형성이나타나서, 인류의각인종을구별할수있는다형성 18
30 또는하플로그룹이존재한다. 또한체세포돌연변이나유전적으로물려받은변이로서각개체를구별해주는다형성이되기도한다. (Brown WM, et al., 1982, Cann RL, et al, 1987, Coble MD, et al, 2004, Scheffler IE, 2008) 여러인종을대상으로미토콘드리아 DNA 염기서열을분석했던 1987년연구에서, 염기서열의해독정보자체는논란의여지가없는것이었으나, 그해석은쉽지않았다. 처음에는단순히돌연변이의숫자만을비교했었다. 일정한곳에정착하여오랫동안거주한인구에서는이인구의표본에서미토콘드리아 DNA의다양성이나타날것으로보았다. 다시말하면, 시간이흐르고세대가교체하면서나타나는뉴클레오티드변화즉분자생물학적시계는계속분침을다투어가며흘러갈것이다. 반대로만약최근에정착하기시작한인구라면미토콘드리아염기서열은변이가거의없는, 비교적동질성으로나타날것이다. 이러한설명은실제로일어나는현상으로밝혀졌다. 이는아프리카에서미토콘드리아 DNA가전해왔을것이라고볼수있는근거로, 이에부합하여아프리카에서현재까지는가장큰유전적다양성을보여주었다. 아프리카인에서관찰되는변이의전체숫자를감안하여추정되는분자생물학적시계로서의개념을근거로한것이었다. (Brown WM, et al., 1982, Cann RL, et al, 1987, Coble MD, et al, 2004) 미토콘드리아 DNA 와인류의계통발생나무 처음변이에근거한분자생물학적접근방법이제시되었을때에는제한효소기법의특성상상세한분석이어렵고한계가있는점, 표본의크기가작은경우정보가간과되거나잘못해석될수있는여지가있는점, 아프리칸미국인이아프리카를대표한다는것은치우침 ( 편견, bias) 이있을수있는점, 계통발생학적나무 (phylogenetic tree) 의개념을수립한방법론에대한의문이제기된점등여러가지비판과논란이있었다. 그러나시간이흐르고기술이발달하면서이렇게제기되었던이러한논란들은더이상문제가되지않았는데, 이를테면제한효소기법의경우더많은종류의제한효소를추가하여분석함으로서분석방법을개선시키거나, 전체염기서열분석을시행하여기존의염기서열분석기법의제한점을극복할수있었고, 다른연구를통하여여러지역의대표인구에대한표본의크기와수를늘려서 19
31 진행하면극복할수있었다. 이러한연구들에서주로선호되는부위는 DNA 복사와전사가시작되는부위가포함되어있는조절부위로, 이부분이가장빠르게진화과정을반영하기때문이었다. 가장적절한계통발생학나무를제시하기위하여 Phylogenetic Analysis Using Parcimony (PAUP) 라고명명된새로운컴퓨터프로그램이개발되었고, 이전의여러연구를통해얻어진막대한양의정보를이프로그램을이용하여분석하였다. 1987년발표되었던여러인종을대상으로분석했던연구자료를이프로그램을통하여 1991년에다시분석하였다. 미토콘드리아 DNA로인류의최초조상은약 200,000여년전에아프리카에살았을것이라고하였다. 다만당시컴퓨터나프로그램은기술적으로발전단계초기로적절한계통을잘찾아내지못하였고, 계통발생학나무의뿌리가정의되지않아서불완전하였다. (Cann RL, et al, 1987, Coble MD, et al, 2004, Vigilant L, et al., 1991) 1996년일명 장식적전정법 (topiary pruning) 이라고불리는방법으로앞서연구를통해알려진자료들을다시분석하였다. 이방법은상기명명된이름이가리키는것처럼애매한새자료를삭제하고확실한계통을보여주도록정리하려는시도였다. 계통발생학적나무역시실제나무가성장하는것처럼서로다른계통이있고각계통의염기서열중돌연변이가잘일어나는부위에서최근에반복적으로돌연변이가형성된다면, 이는기존나뭇가지에서자라나는 새가지 가될수있을것이다. 또한이러한 새가지 의계통이더중요한기존가지인계통을가려버릴수있다는점에서, 가지치기가필요하다고보았다. 이렇게정리된나무는계통분류의가짓수를줄여주고단순화시키면, 여러통계학적방법에의한분석 ( 가지치기, 가중치주기, 자동처리등 ) 에적합하게된다. 진짜계통발생학적나무에가까워질수있기위해서는이러한여러통계학적방법의복합적인조합에의해가능하다고보았기때문이었다. 이접근방식으로분석해본결과, 인류의미토콘드리아유전의기원이아프리카임이직접적으로증명되지는않았으나, 가지가정리되어얻어진계통발생학적나무에대한분석결과는아프리카가기원의가능성에무게를두었다. (Scheffler IE, 2008) 시간척도의측정에대해서도논란이있었다. 변이가관찰된모든부위에서다같은속도로변화가일어나는것인가? 조절부위가암호부위보다더빠른속도의변화를보이는가? 코돈의세번째위치에 20
32 나타나는변이에의한변화가더빠른속도로발생하는가? DNA 염기서열에서전이 (transition) 와변이 (transversion) 의발생속도를서로비교하면어떻게되는가? 이러한의문들을맞닥뜨려인류미토콘드리아 DNA 변이의더정확한시계척도를예측하기위한연구가시도되었다. 아프리카에서기원한현생인류는 년전에출현하였을것으로추정할수있었는데이분석결과는현생인류의기원은한지역이아닌여러지역에서왔을것이라는정면으로배치되었다. 미토콘드리아 DNA에발생하는돌연변이의발생과그변화의속도에관련된이러한불확실성과논란은모든관련전문가들의눈높이를만족시킬수없기때문에미토콘드리아 DNA의최초조상의나이에대한추산은 10만년이하에서부터거의 100만년전으로까지광범위하였다. 미토콘드리아 DNA 염기서열로수립된계통발생학적나무를둘러싼이러한논란은유전자관점에서보는계통과각개체의관점에서보는계통을구별하지않음에서비롯된것으로다양한인종의미토콘드리아 DNA를분석하여얻어진하나의조상염기서열 ( 미토콘드리아이브 ) 이모든현생인류가하나의모체에서나왔다는것을의미하는것은아니기때문이다. 하나의모체가아니라면현생인류가형성된초기인류의규모는어떠하였는가에대한의문은여전히남아있었다. 미토콘드리아 DNA의계통발생학적뿌리는아프리카였을가능성은있지만, 초기인류에서병목현상의존재유무및존재한다면그규모, 생존한여러인류의혼합에따른복합병목현상등에대한의문이남아있었다. 그래서인류와가까우면서도더조상격인다른종에대한연구도시도되었다. (Ayala FJ, 1995, Gibbons A, 1992, Hasegawa M, et al, 1993, Scheffler IE, 2008) 미토콘드리아 DNA 와계통발생학나무를통한인류의기원 연구와 DB 구축 여러연구에의해진행된미토콘드리아 DNA 염기서열분석정보를종합하여분석한결과지구에형성된인류에대한가설을다음과같이제시하였다. 10만에서 20만년전무렵아프리카에서인류는이주하기시작하였고, 약 5만년전에한군의인류가중동지역에서다시유럽중부와북부로갈라져나왔으며, 또다른군이아시아남쪽과동남쪽으로이동하였다. 또한오래전에중국지역에서인도네시아 21
33 섬지역을거쳐뉴기니아, 호주로이주하였다. 이후이인구는아시아동쪽끝을따라북쪽으로확장하여베링해협을거쳐북미에이르렀다. 이후알래스카에서남쪽으로진행하여북미, 중미, 남미로확장하여나갔다. 뉴기니아와오스트레일리아북쪽주면의섬에서북태평양 ( 폴리네시아, 하와이등 ) 으로인구가확장하여이주한것이가장최근의인류의역사로보았다. 이러한가설은고인류학, 고고학, 비교언어학에서도인류의역사를유사하게설명하고있어서완전히새로운이론이라고할수는없었으나미토콘드리아 DNA 염기서열분석정보는좀더상세하면서확실한근거를제시할수있었다. 그리하여인류의계통발생학적나무의뿌리는아프리카에있다는점은어느정도과학적합의가이루어지게되었다. (Scheffler IE, 2008, Redd AG, et al., 1995, Wallace DC, 1995) 인류의기원에대한지역은합의가되었으나미토콘드리아 DNA의조상인이브의나이에대해서는여전히논란이분분하였다. 지구상의여러지역에서발견된화석에서제시하는근거 ( 직립원인은적어도백만년이전존재 ) 를감안하였을때현생인류가탐지가능한유전적교환이없이이전의인종을완전히대체하였을것이라는유전학적근거에의거하여인류의기원을설명할수밖에없을것이다. 미토콘드리아이브의나이가약 50만년이상이라면다지역발생설, 인체유전자의구성의변화, 특이한진화유형을형성하도록하는선택적압력등이러한점들을둘러싼여러가설이제기될수있었다. 1995년에기존에알려진분석자료를다시철저하게재분석하였고미토콘드리아이브의나이는아마도 43만6천년에서 80만6천년사이일것으로보았고, 이러한추정은인간과침팬지의계통사이에서갈라지는시간대를추산하여근거한것이었다. 이후미토콘드리아 DNA외에도 Y염색체, 상염색체등여러인간 DNA 염기서열분석정보가밝혀지면서 2006년새로운설명이제시되었다. 인류의역사는크게두시대로나뉘는데첫번째시대는약 2백만년전에나타났으며현생인류와해부학적으로유사하였다. 두번째시대는아프리카에서현생인류가출현하여지구상의각지역으로분포된다. 염기서열분석은유전자나무를수립하는데사용되었고이나무는뉴클레오티드변이가축적이분자생물학적시계가될수있는가정에근거하여가장최근의조상의시간을추산하는데사용되었다. 이분석결과를종합하여보았을때, 인류의조상은 50만년에서 80만년으로추정되며현생인류이전에소멸된네안데르탈인등여러계통을포함하는것으로단일기원설보다는 22
34 다지역기원설에기울어지는가설이었다. (Scheffler IE, 2008, Wills C, 1995). 이렇게여러인구에대한연구를통해확인된미토콘드리아 DNA 염기서열과다형태의정보들을등록하고공용으로이용할수있는미토콘드리아 DNA 데이터베이스 ( 가구축되어있다. 이외에이형세포질성에대해검출된자료의신뢰성을확인해볼수있는 mtdna-server ( 등이있다. (Jin HJ, et al., 2009, Lee HY, et al., 2006, Weissensteiner H, et al., 2016, 그림2 & 그림3 참조 ) 미토콘드리아 DNA 와하플로그룹 하플로그룹에대한개념은미주원주민을대상으로한미토콘드리아 DNA에대한연구에서처음제시되었다. 조절부위염기서열을분석한결과관찰된변이의양상에따라 4개의하플로그룹 ( 하플로그룹 A, B, C, D) 으로분류하였고, 이후발견된하플로그룹에대하여다른알파벳을사용하여분류하였다. 1998년서유럽의여러인종에대한미토콘드리아 DNA 조절부위와암호부위일부특정부위를바탕으로분석결과확인된유전적다양성을하플로그룹으로명명하는법을제시하였다. 그러나이후여러연구들에의해이루어진미토콘드리아 DNA 하플로그룹명명이일치하지않는경우가있어서, 같은하플로그룹을다른이름으로명명하거나서로다른하플로그룹이같은이름으로명명되곤하였다. 이러한불일치는모두가합의할수있는일반적이면서체계적인하플로그룹분류시스템의필요성이대두되었다. (Richards MB, et al., 1998, Torroni A, et al., 1993, van Oven M, et al., 2009) 2009년전세계인류의미토콘드리아 DNA 다양성을보여주는수많은염기서열변이들에대해서손쉽게이용할수있도록하기위하여관련여러문헌들을검토후포괄적으로종합하여정리한계통발생나무및하플로그룹분류시스템이제시되었으며이는기존에알려진개정캠브리지표준염기서열 (rcrs) 을바탕으로한것이었다. 이후이개정캠브리지표준염기서열은유럽인의하플로그룹의하나인 H2a2a1에속한것임이알려지면서좀더인류미토콘드리아 DNA의계통발생학적인계보를잘보여줄수있는개정된인류표준염기서열 (Reconstructed Sapiens Reference Sequence) 이제시되었다. 다만이를실무에바로적용하기에는아직불완전하기때문에개정캠브리지 23
35 표준염기서열을표준으로사용하는것이권고되고있다. 이러한표준염기서열을바탕으로여러연구를통하여미토콘드리아 DNA 염기서열전체에서확인된변이의양상에따라계통발생학나무에따른다양한군의하플로그룹으로분류하고각유형에는영어알파벳과숫자를이용하여명명하였다. 모든하플로그룹은계통별로정리되어 Phylotree ( 에종합되어있으며, 여러연구를통해보고되는새로운변이를지속적으로갱신하여반영하고있다. ( 그림 3 참조 ) 또한인류에대한진화인류학, 인류역사, 의학유전학, 유전계통학, 법의학등의연구를위한도구로사용할수있기위하여구축된것으로주기적으로갱신되고있다. 또한이계통발생나무를근간으로, 하플로그룹을손쉽게결정할수있는온라인분석프로그램 (MitoTool, HaploGrep2, 이개발되었다. 이러한데이터베이스에수록된자료들을근거로하플로그룹을결정할수있고, 이는계통발생학적의의뿐만아니라법의학개인식별영역에서개체간식별에사용되는과학적근거가되었다. 그밖에도하플로그룹으로특징지어지는다양한변이의특성은미토콘드리아기능에영향을주고특정질병과의관련성이있음이알려지면서하플로그룹의분류는개체식별뿐만아니라질병의연구에서도중요한의의가있다. (Behar DM, et al., 2012, Hasegawa M, et al., 1993, Scheffler IE, 2008, van Oven M, et al., 2009, Samuels DC, et al., 2006, Stewart JB, et al., 2015, Weissensteiner H, et al., 2017a, Weissensteiner H, et al., 2016b, 그림3 참조 ) 2.3. 미토콘드리아 DNA 의법의학적활용의역사 미토콘드리아 DNA 를이용한개인식별의실제적인사례 1983년코카서스인과아프리카인을대상으로작은규모로선별하여진행한연구에서미토콘드리아 DNA 조절부위중 2곳의과변이부위에집중적으로뉴클레오티드변이가몰려있음이발견되었고, 통계적분석을통해개인식별에적용하였다. 과학기술의발전으로사법절차와인권보호를위한활동에과학이근사하면서도감동적인모습으로 24
36 접목되었던계기는 1983년아르헨티나정부의요청에따른미주인권위원회의활동이었다. 아이들의엄마들이군사정권하에희생되어당시생존한아이들이다른곳으로입양되었는데, 아르헨티나법정에서는이아이들의친자관계가과학적으로증명될수있다면헤어졌던가족과연결해주려고하였다. 친부모가없었던상황에서법적으로친자관계를증명할수있었던방법은모성유전인미토콘드리아 DNA 염기서열분석을통해아이의하플로그룹과예상되는외할머니친족들의하플로그룹과비교하는것이었다. 다만이방법이과학적으로는객관적이고결정적이었지라도, 여러법적, 윤리적, 사회학적, 심리학적쟁점들이함께부상하였다. (Anderson S, et al., 1981, Scheffler IE, 2008, Greenberg BD, et al., 1983) 미토콘드리아 DNA 를이용한개인식별과이형세포질성 미토콘드리아 DNA 염기서열분석을적용했던또하나의놀라운이야기는러시아의황제였던니콜라스 II세와그의부인, 아이들, 그들의종복들의유해에대한진위여부에대한것이었다. 러시아의황족들은혁명기동안처형되어예카테린부르크근방에서공동묘지에묻혔다. 1991년 9구의유골이발견되었고여기에황제와그의황족들이포함되었는지증명이필요하였다. 뼈조직에서미토콘드리아조절부위의염기서열이확보되었기때문에그들의생존해있는친척과사망한친척들과의대조하였으며, 이때미토콘드리아 DNA 조절부위의점이형세포질성에대한후속연구들의시발점이되었다. 미토콘드리아 DNA 다형태라는특징을사법절차에적용한다는개념은미국연방수사국과군대가각별한관심을보였다. 120이상의가족을대상으로미토콘드리아조절부위염기서열을분석하였고이결과로인하여 호발부위 (hot spot) 의가능성, 돌연변이발생율과진화의속도및인류의진화에대한논란이가열되기도하였다. (Scheffler IE, 2008) 한국에서미토콘드리아 DNA 와개인식별연구 2002 년미토콘드리아조절부위염기서열이외에암호부위중시토크롬 B 유전자의다형태를한국인에서개인식별에적용한연구가발표되었다. 25
37 이후한국인에서미토콘드리아조절부위의이형세포질성에대한연구, 한국인에서나타나는미토콘드리아 DNA 조절부위를중심으로다형성과하플로그룹, 이형세포질성양상을파악하고자하는연구를통해한국인의개인식별에이를활용하였다. (Choung CM, et al., 2005, Holland MM, et al., 1999, Hong SB, et al., 2014, Jin HJ, et al., 2009, Lee HY, et al., 2006, Lee SD, et al., 2002) 3. 법의학에서미토콘드리아 DNA 최근연구동향 3.1. 법의학에서미토콘드리아 DNA 에대한기존연구의한계 미토콘드리아 DNA 분석의한계 : 제한적인개체식별력 오래된유골이나머리카락처럼세포핵 DNA가변성되거나분석이불가능한경우미토콘드리아 DNA를통해모계와비교분석, 미토콘드리아 DNA에서확인되는변이의양상을통해결정된하플로그룹으로개체간식별을위한추가적인정보를확보할수있는점은미토콘드리아 DNA 분석의강점이었다. 그러나이러한강점은동시에약점이되기도하는데세포핵 DNA만큼강력한개체식별력을주지못하기때문이다. 특정지역의인구내에서하플로그룹의분포는비대칭적인분포를보이기때문에흔한유형의하플로그룹으로결정되는경우에는개체식별의한계에부딪히게된다. 예를들면코카시안미국인을대상으로한연구에서약 20% 의개체가인구의 0.5% 이상의빈도를가지는미토콘드리아 DNA 하플로그룹을가지고있다고하였다. 즉동일모계가아님에도우연히같은하플로그룹을가질가능성이있으며, 이는개체간식별의제한점이되었다. (Coble MD, et al., 2004, Fendt L, et al., 2009, Holland MM, et al., 1999, Just RS, et al., 2015, Lee HY, et al., 2006, Lee SD, et al., 2002, Nilsson M, et al., 2008, Parsons TJ, et al., 2001) 기존미토콘드리아 DNA 분석의한계 : 분석기법측면 미토콘드리아 DNA 에대한기존의연구와실무는생거의 26
38 염기서열분석기법이주로적용되었으며, 미토콘드리아조절부위에서변이가자주나타나는과변이부위및암호부위일부특정부위가실무적으로는주분석대상이었다. 이는연구기법적인면에서시간, 비용과인력의소모를최소화할수있는방법이었기때문이었다. 개체간식별력을높이기위하여기존의미토콘드리아 DNA 조절부위의과변이부위를벗어나암호부위등다른부위에서관찰되는변이를포함하려는연구가우리나라를포함하여진행되었다. 이들연구를통하여미토콘드리아 DNA 조절부위뿐만아니라암호부위에대한염기서열분석을통해식별력이증가될수있음이어느정도증명되었다. 2009년에는이러한제한점을감안하여조절부위와암호부위전체를포함하는전체염기서열분석을수작업을통해높은질의결과를보여줄수있는방법을제시하였었다. 다만현실적으로전체염기서열분석은여전히비용효율적인면에서실무에적용하기에는어려운상황이었다. (Coble MD, et al., 2004, Fendt L, et al., 2009, Holland MM, et al., 1999, Just RS, et al., 2015, Lee HY, et al., 2006, Lee SD, et al., 2002, Lutz-Bonengel S, et al., 2003, Nilsson M, et al., 2008, Parsons TJ, et al., 2001) 3.2. 차세대염기서열분석기법의도입 새로운 DNA 분석기법의필요성대두 1990년대에서부터 DNA 염기서열분석기법은모세관기반으로반자동으로수행되는생거기법에의해수행되었다. 대량의염기서열분석을수행하는경우 2가지방식으로분석한다. 첫째는샷건염기서열분석기법 (shotgun de novo sequencing) 은 DNA를무작위로잘게토막을낸후에이를대장균을변화시킬때사용되는플라스미드에주입하여많은복제수로복제시키고플라스미드 DNA만을추출하여염기서열을해독하는방법이고둘째는해독하고싶은관심부위의염기서열에상보적으로일치하는길잡이를통해복제하여염기서열을해독하는방법이다. 염기서열분석은 주기염기서열해독 (cycle sequencing) 반응으로형성되며각주기에서는 DNA 주형의변성, 길잡이결합및연장등이일어나고, 길잡이는 27
39 관심부위에상보적으로결합하는염기서열을가지고있다. 길잡이에서염기서열이연장될때, 형광물질표지자가붙은디디옥시뉴클레오티드가확률적으로결합하면, 염기서열의연장은중단되고, 각 DNA 토막은중단된부위에표지자가붙어있게된다. 모세관기반의중합체겔에이들표지자가붙어있는 DNA 토막들을정주하면고해상도전기영동방식으로한가닥의표지가붙은 DNA로분리가되면서염기서열을해독할수있게된다. 레이저를통해형광물질표지자를자극하면, 4가지색깔을추적하여인지하고, 이를컴퓨터소프트웨어를통해추적한결과를염기서열로해독한다. 다만각염기서열검출에는오류가있을확률도있다. 약 30년동안점진적인기술개발로생거기법은 1000개의염기쌍까지해독할수있으며, 염기서열검출의정확도는 % 로매우높다. 대량의염기서열을분석할수있는샷건염기서열분석기법의경우매 1000염기쌍을해독하는데미화로약 0.5불의비용이소모된다. (Lutz-Bonengel S, et al., 2003, Metzker ML, 2010, Schendure J, et al., 2008) 인간게놈프로젝트가진행된이후기존의 DNA염기서열분석기법의비용을감소시킬수있는적절한다른길이거의남아있지않은점, 인간과주요생명체의전체유전자의표준서열이있어서이는짧은길이의 DNA 염기서열해독이잠재적으로매우유용할수있는점, 분자생물학적기법이매우다양하게발전하면서광범위한생물학적현상이대량의 DNA 염기서열분석으로평가되고있다는점, 마지막으로현미경, 표면화학, 뉴클레오티드생화학, 중합체공학, 컴퓨터공학, 데이터저장및기타영역등서로다른영역에서전반적으로기술이발달하면서, DNA 염기서열분석기법에대한대안기법이실제적으로사용될수있도록기여하는점등이러한 4가지면에서새로운차세대염기서열분석기법의필요성이대두되었다. (Metzker ML, 2010, Schendure J, et al., 2008, Shendure J, et al., 2004) 생거방법의기술적변화 : 피로시퀀싱 차세대염기서열분석중간과도기무렵에기존의생거방법을변화시켜 개발된 방법이 있었다. 피로시퀀싱 (pyrosequencing) 이라고 하며, 전기영동으로 보여주기 전에는 방사성 또는 형광표지자가 붙은 뉴클레오티드 또는 올리고핵산염 (oligonucleotides) 을 사용하여 뉴클레오티드 서열을 추정할 수 없다는 점에서 기존의 방법과 다른 28
40 점이다. 피로인산염 (pyrophosphate) 이합성되면이반응으로빛을발하는현상을이용하는방법이발견되어이를이용한것이다. 이는 2개의효소로구성된것으로 DNA 주형에상보적염기서열이연장될때피로인산염이방출되고, ATP sulfurylase가이피로인산염을 ATP로변환시키고, 이는루시페레이즈의기질로작용하여피로인산염의양에따라빛이나오는발광시스템을적용하였다. 고체상에부착된 DNA주형에차례로뉴클레오티드가분주될때마다형성되는피로인산염의양에따라염기서열을추정하게된다. 이방법역시기존의생거방법처럼 sequence-by-synthesis, 즉적절한결과를얻기위해서는 DNA 중합효소의직접반응이필요하다는점에서는다르지않았지만, 디디옥시뉴클레오티드를사용하지않고일반적인뉴클레오티드를사용해도되는간편한점이있고전기영동이필요없이실시간으로관찰할수있다는장점이있었다. 후에 DNA 주형을상자성구 (paramagnetic beads) 에부착시키고, 효소작용으로변성된뉴클레오티드를제거하는등방법을개선시켰다. 이방법의어려운점은같은뉴클레오티드가연속적으로배열된염기서열의경우몇개까지동일뉴클레오티드인지알아내는것으로, 5개이상인경우해독이어려워진다는것이었다. 이방법이더진화되어차세대염기서열기법의첫성공모델이되었다. (Heather JM, et al., 2016) 차세대염기서열분서기법의탄생 : 초병렬염기서열분석방식 454 (Life Science) 는기존의인식체계의변화를가져온염기서열분석기계로염기서열분석반응을초병렬로수행하여한번의분석으로수행할수있는 DNA의양을증가시킬수있었다. 유도자 (adapter) 의염기상보결합을통해 DNA를구에붙이고이른기름속에물형태의유탁액 PCR을진행시키면, 유탁액안의각방울에는이상적으로한종류의 DNA로동일하게복제된상태로구가덮이게된다. 이후이를세정하여피로시퀀싱방식으로효소반응을시키고광전변환소자 (charged couple device) 감지기를통해염기서열을해독하게된다. 이방법은보통길이 염기쌍을해독할수있었다. 대량의 DNA를병렬식으로염기서열분석을미세수준에서수행할수있다는것은미세수준의제작과고해상도영상기술덕택이었으며이른바제 2세대 DNA 염기서열분석기법또는차세대염기서열분석기법이새로운장을열게되었다. (Heather JM, et 29
41 al., 2016) 제2세대 염기서열분석기법은 몇 가지로 나뉠 수 있는데, 미세전기영동방법, 부합화 (hybridization) 을통해염기서열을해독하는 방법, 각 입자를 실시간으로 관찰하는 방법, 주기적인 배열 (cyclicarray) 로 염기서열을 해독하는 방법 등이 제시되었다. 주기적 배열 염기서열분석기법 (cyclic-array sequencing) 이 주로 여러 가지로 사용되었다. 이기법은샷건염기서열방식과주기적배열방법을결합한 기법이다. DNA를무작위로여러토막으로쪼갠후에동일한염기서열의 유도자 (adapter, 짧은 염기서열 ) 와 DNA 토막들을 결합시켜 DNA 라이브러리를준비시킨다. 복제된엠프리콘들이군집을이룰수있도록 하기위해서 in situ polonies (PCR colony), emulsion PCR, bridge PCR 등여러가지프로토콜이사용될수있다. in situ polonies 방식으로 진행시키는 경우 수백만의 공간적으로 움직일 수 없는 폴로니의배열들이나타난다. 각폴로니에는한개의샷건 DNA 토막의 많은복제수가있다. 한번의마이크로리터의시약양 ( 길잡이결합후 염기서열 연장반응 ) 으로 모든 배열을 병행하여 다룰 수 있다. DNA형광물질 표지자를 인지하여 영상으로 변환하고 염기서열로 해독하며, 이모든과정은병행적으로이루어진다. bridge PCR 은 유도자와 결합된 DNA를 flow-cell 에 부착된 상보적 올리고핵산염에 정주하고 고체상 PCR을 수행하면 각 flow-cell에 결합한 DNA 주형으로부터동일한복제군집이나타난다. 처음결합했던 올리고핵산염에서염기서열이연장되어바로이웃한올리고핵산염으로 연결되면서고리모양으로형성되기때문에 bridge 라고명명되었다. 이경우 DNA중합반응에따른염기서열연장은생거방식을이용하였다. 2005년에 이들 기법이 기전 방법에 비해 실용적이면서도 비용에서 효율적인 방법으로 기술하였다. 기존 생거방법에 비해 전반적인 장점으로는, DNA 라이브러리가 준비되고 이를 복제시킬 때, 기존 방법의 병렬진행을 막는 여러 병목현상을 피할 수 있는 점, DNA염기서열을해독함에있어훨씬더높게병행적으로이루어지는 장점이있는점, 평면의표면에고정되어있어서한번의시약주입으로 효소반응관리가가능하다는점등이있었다. 사용되는시료의양을줄 일수 있었던 점이 비용을 극적으로 감소시키는데 결정적인 역할을 하였다. 그럼에도 여러 단점이 이러한 장점들을 상쇄시켰는데, 기존방법에 비해 해독할 수 있는 염기서열의 길이가 훨씬 짧고, 생거방식으로알게된염기검출보다 10배이상정확도가좋지않았기 30
42 때문이었다. 이러한제한점들은곧알고리듬으로중요하게해결해야할 도전이되었다. (Metzker ML, 2010, Schendure J, et al., 2008) 차세대염기서열분서기법 : Ion Torrent PGM 방식 여러기술적인혁신속에서도방사성이든, 형광이든, 발광시스템이든, 빛을감지하여영상을통해해독하는점은여전히동일하였다. 이러한개념을탈피하여인식체계의변환을가져온혁신적인기술이개발되었는데, 이른바 post-light sequencing 으로불리기도하는 2011년에첫선을보인 Ion Torrent Personal Genome Machine (PGM) 이다. DNA 염기서열분석결과를특수뉴클레오티드나특수시약을사용하여빛을이용한이미지로보여주는기존의방식에따른한계를극복하고 DNA 분자그자체를그대로인지하여분석하는염기서열분석기법을개발하려는시도끝에탄생한기법이다. 신개발직접회로를적용하여비시각적인방법으로염기서열분석결과를보여주는기법이었다. DNA 중합효소반응으로뉴클레오티드가결합하는과정에서수소이온이방출되고, 이로인해 ph가변하게된다. 시모스반도체를이용한이온칩을통하여이러한 ph변화를트랜지스터의전위변화로인지함으로서염기서열분석이가능하게되며, 이를통해동시다발적으로대량의 DNA 중합효소반응을인지한다. 상보성금속산화막반도체즉시모스 (Complementary metal oxide semiconductor; CMOS) 라는직접회로의한종류를응용한것으로, 널리사용되는기술을채택함으로더낮은비용으로대량의정보를더빠르게처리할수있도록한것이이분석기법의장점으로꼽힌다. DNA 처리방식은피로시퀀싱과유사하게구에동일하게복제된다수 DNA 토막들을 picowell에세정시키면각뉴클레오티드가결합하여염기서열이연장될때이를피로인산염이아닌 ph의변화로감지한다. 이모든과정은작은칩에서일어나며추가적인광학장비없이바로컴퓨터소프트웨어에서처리하여결과를보여줄수있게되었다. 이러한기술적인새로운변혁은보편화된기술을도입하여비용을낮추면서도염기서열해독의감지도를높임으로인간의유전체염기서열해독을보다손쉽게접근할수있게해주었고, 미토콘드리아 DNA 염기서열분석에도이기법을적용하기시작하였다. (Quail MA, et al., 2012, Rothberg JM, et al., 2011) 31
43 3.3. 법의학에서차세대염기서열분석기법을적용한미토콘드리아 DNA 최신연구 차세대염기서열분석기법을적용한미토콘드리아 DNA 염기서열분석기법의입증과수립 2013년차세대염기서열분석기법인 Ion Torrent PGM을적용하여미토콘드리아 DNA의전체염기서열분석을시도하고이를전통적인방법인생거기법으로분석한결과를비교분석함으로서, 실무영역에서이기법을시도해볼수있는기틀을마련하였다. 혈액, 파라핀블럭의조직, 구강점막에서면봉으로채취한시료등여러시료를대상으로하였으며생거기법을적용한염기서열분석도함께시행하였으나일부시료는이전발표자료를일부포함하였다. 과변이부위에위치한 3곳의 C뻗기부분, 이외의결실, 치환, 점이형세포질성에대해서비교분석, 정렬알고리즘으로비교분석등을시행하였다. 분석결과대부분의염기서열이일치함을알수있었으며, 불일치로보였던염기서열분석결과도염기서열분석과정의정렬알고리즘과관련된것으로생각되었다. 이연구결과는차세대염기서열분석기법을법의학실무영역에서적용하였을때, 정렬알고리즘의개선, 데이터분석과해석에대한새로운지침등의필요성을제시하는근거가되었다. (Fendt L, et al., 2009, King JL, et al., 2014, Kong QP, et al., 2003, Parson W, et al., 2013, Seo SB, et al., 2015, Stoljarova M, et al., 2016, Templeton JE, et al., 2013, Zhou Y, et al., 2016) 차세대염기서열분석기법에의한다양한인종에서의 미토콘드리아 DNA 전체염기서열분석및유전적다양성연구 이후 차세대연기서열분석기법을 적용하여 여러 인종에서 나타나는 미토콘드리아 DNA 전체염기서열을해독하여분석한연구들이뒤를 이었다. 2014년 미국의 코카시안, 히스패닉, 아프리칸 미국인을 대상으로미토콘드리아 DNA 전체염기서열분석및하플로그룹에대해 보고되었으나, 이는 기존 생거 방식을 따른 것이었다. 2015년 에스토니안 인종에 대하여 차세대염기서열분석기법을 적용하여 미토콘드리아 전체염기서열분석을 시행하였으며 분석결과, 기존의 32
44 미토콘드리아조절부위과변위부위로만하면 95.85% 의유전적다양성을보여주지만, 전체염기서열을분석하여보면 % 의유전적다양성을보여주었다고하였다. 2015년모발일부분이나빠진모발처럼털집이없는모발에서미토콘드리아 DNA 정보를얻기위해초병렬염기서열분석기법을이용하여전체염기서열분석결과를얻어내어법의학업무에서차세대염기서열분석의유용성을증명하였다. (Fendt L, et al., 2009, King JL, et al., 2014, Kong QP, et al., 2003, Parson W, et al., 2013, Seo SB, et al., 2015, Stoljarova M, et al., 2016, Templeton JE, et al., 2013, Zhou Y, et al., 2016) 미토콘드리아 DNA 전체염기서열분석도구의변화 미토콘드리아 DNA 분석에 차세대염기서열분석기법의 도입으로 전체염기서열에 더 쉽게 접근할 수 있게 되었으나 그 결과 얻어진 막대한 양의 정보의 처리, 민감도가 높아지면서 실제 돌연변이와 염기서열분석과정의인공산물을구분하는등의해석등기존과는다른 분석도구의 필요성이 대두되었다. 차세대염기서열분석기법을 통해 얻어진 염기서열정보를 입력하면 자동화된 온라인 프로그램에서 분석하여 결과를 제시해주는 MitoTool, HaploGrep 등의 분석도구가 탄생하였다. 개정캠브리지표준염기서열 (rcrs), 개정인류표준염기서열 (RSRS) 에따른계통발생학적분류에따라분류된 하플로그룹에 대한 결과가 제시된다. MitoTool 은 생물정보학에 근거한분석도구로, 계통발생나무에따라하플로그룹을결정할때각 하플로그룹을결정하는주요변이와분석하고자하는시료에서나타나는 변이들의공통점과차이점을분석하여점수를주는방식으로접근한다. HaploGrep 은하플로그룹을결정하는데가장정확한알고리즘으로 분석하는 것으로 알려져 있으며, MitoTool 과의 차이점은 HaploGrep 은 각 변이에 대하여 계통발생학적으로 얼마나 자주 나타냐는가 하는 빈도를 근거로 가중치를 부여하여 분석하고, MitoTool 은 이런 가중치를 부여하는 점이 없는 것이 차이이다. 따라서 개별변이 (private variant) 가 있는 경우 가중치 부여에 따른 영향으로그의의가무시되거나또는더해질수있는치우침 (bias) 이 있을 수 있다는 의견도 제시되었다. 또한 차세대염기서열분석기법의 특성상 적은 양의 DNA 염기서열분석도 가능해짐에 따라 33
45 이형세포질성의결정과해석이더복잡해졌다. 그리하여이형세포질성분석만을위한웹분석도구가개발되었는데바로 mtdna- Server 이다. 차세대염기서열분석결과를입력하여, 분석결과의질을평가한후에, rcrs에서변이호발부위 (hot spot; 309, 315, 3107 등 ), 10 미만의낮은 coverage를보이는부위를포함시키지않으며, 여러알고리즘을통해분석하고 NUMT( 세포핵내미토콘드리아염기서열 ) 에대해서분석하고, 시료간오염의가능성도검토할수있게되었다. (Fan L, et al., 2013, Weissensteiner H, et al., 2016) 3.4. 동아시아인종및한국인의미토콘드리아 DNA 대한연구 2002년 중국 한족에 대한 미토콘드리아 DNA를 분석하여 계통지리적인 분석을 시도하였다. 중국 한족은 세계에서 가장 큰 인종으로전체지구인구의 20% 를차지한다. 이전에도중국인에대한 연구가있었고, 같은해중국남쪽지역의 69명의중국인을대상으로 미토콘드리아전체염기서열분석을한다른연구가있기는하였으나, 중국대륙전체인구와분포를고려하였을때, 시료선별이제한적이라고 보아 중국의 6개 지역에서 263명을 선별하여 이들의 미토콘드리아 DNA 조절부위의 과변이부위 염기서열을 분석하였다. 각 지역별로 나타난하플로그룹은달랐으며, 확인된하플로그룹으로는 M7b1, M7b2, M7b, M7c, M7, M8a, C, Z, M9, M10, M, N, M/N, G2, D, D4a, D4b, D4, D5a, D5, A, N9a, Y, B4a, B4b, B4, B5a, B5b, B5, B, R9a, R, F1a, F1b, F1c, F2a, F2, F 등이 있었고 일부 서유럽 인종으로 알려진 하플로그룹도있었다고하였다. (Kivisild T, et al., 2002, Underhill PA, et al. 2007, Yao YG, et al., 2002) 다음해인 2003년에는한족이외에중국북쪽지방에거주하는 5군의 인종 ( 다우르족, 에벤키족, 조선족, 몽고, 어룬춘족 ) 의미토콘드리아 DNA 조절부위의 과변이부위와 암호부위 일부 대하여 염기서열분석을 시행하였다. 각인종에따라공통적으로보이는하플로그룹 (D, D5, A) 도있었지만대부분은각인종에따라다른유형을보였다. 이주한 한국인으로생각되는조선족의경우공통유형이외에하플로그룹 A5, G2, G2a, Z, M10, M7b2, B4b, B5b, G1a, G3, M7a1, M7c, M, N9a, B4 등이있었다고하였다. (Kong QP, et al., 2003) 이렇게 발표된 여러 연구에서 일부 새로 정의된 하플로그룹을 34
46 반영하여제시한동아시아계통발생학적나무가만약미토콘드리아 DNA 전체염기서열분석결과에부합할만큼신뢰할만하고동아시아의미토콘드리아 DNA의가계의관계를완전히반영하는지에대한의문이제기되었다. 같은해중국전역에서시행된 2000여개이상의샘플중에서서로다른하위하플로그룹을지닌 48개의시료를선택하여전체염기서열분석을시행하였다. 하플로그룹 D, M7, C, A, N9a, M, R, N 등은부합한결과를보여주었다. 하플로그룹 D5, G1, G2, M7b, M7c, M8, M8a, Z, M9a, M10, F1, F1a, F1c, F2, B4c, B5, B5a, B5b 등은추가적인변이로유형결정이가능하였고, 일부하플로그룹은이연구를통해재정의되었다. (Parson W, et al., 2015) 2004년 672명의일본인을대상으로전체염기서열분석을시행하여이에대하여계통발생학적분석을하였다. 이때의하플로그룹유형결정은초기분류에따른것으로, 미토콘드리아 DNA염기서열분석을통해일본인은대륙아시아에서이주하여왔을것이라고제안하였다. 하플로그룹 A1, N9a이가장흔한빈도를보이며, 이외확인된하플로그룹으로 F1b, B5b2, A1a, N9b, F1a1, B4c1, M/N, B, B5b1, HV, B4a, B4a1, B4, B4c1b, B5a1, B4b, B5b, B4f, F, A1b1, F1a, R9a B5b3 등이있었다. (Tanaka M, et al., 2004) 미토콘드리아 DNA 전체염기서열을분석한연구보다는조절부위와암호부위일부부위를적용한연구들이진행되어왔었다. 2002년미토콘드리아조절부위이외의부위이외에다형성이많은부위로시토크롬B 유전자를적용하여개인식별력을개선하려는연구가있었다. 암호부위의특정부위에해당하는시토크롬B 유전자의다양성은조절부위염기서열이동일한것으로여겨졌던개체군에서세부식별이가능함이증명된연구였다. (van Oven M, et al., 2009, Bandelt HJ, et al, 2001) 2006년 593명의한국인을대상으로미토콘드리아 DNA 조절부위염기서열분석을하였고확인된여러변이에대한데이터베이스가구축되었다. 한국인에서나타난하플로그룹에는 D4, D4a, D4b, D4b1, D4b2, D5, D5a, d5b, D, G1a, G1b, G2a1, G2a1a, G2a2, G3a, M7a, M7a1, M7b1, M7b2, M7c, M7c1, M8, CZ, C, Z, M8a, M9a, M10, M11, M, F F1, F1ac, F1a, F1ac, F1a, F1c, F1b, F2a, R11, B4, B4a, B4b1, B5a, B5b, A, A4, A5, N9a, N9a1, N9a2, N9b, Y, Y1, ND 등으로알려졌고, 이중가장흔한하플로그룹유형은 D4, D4a, B4, B4a, M7b2 등이었다. 같은해대부분의한국인의미토콘드리아 DNA 조절부위에는 35
47 동아시아에서나타나는여러하플로그룹을결정하는중요한변이들을공유하고, 세부하플로그룹을결정하기위해서는암호부위의변이가필요하다는점에서암호부위변이중선별한단일염기다형성표지자를이용하여하플로그룹결정의정확성을높임으로개체식별력을높이기위한연구였다. (Lee HY, et al., 2006, Lee HY, et al., 2006) 2009년한국인에대하여계통발생학적접근을하는연구가있었다. 한국인, 조선족, 몽고, 만주족, 한족, 베트남인, 태국인등동아시아의 7곳의인종에대하여미토콘드리아 DNA 조절부위과변이부위, 암호부위염기서열중하플로그룹유형결정에특이적인변이, Y 염색체변이에대하여분석하였다. 일반적으로한국인의미토콘드리아 DNA는하플로그룹의분포를분석해보았을때동북아시아인종과유사하였고, 일부남아시아인종이변이형성에기여하였을것으로보았다. (Jin HJ, et al., 2009) 4. 연구의필요성 미토콘드리아 DNA의염기서열해독및분석은유전적다양성에대해파악할수있을뿐만아니라법의학의개인식별영역에서기초자료가된다는점에서매우중요하고필요하다. 최근의과학기술의발달로탄생한차세대염기서열분석기법과같은신기술은미토콘드리아 DNA 전체염기서열분석을손쉽게하였고, 법의학개인식별영역에서도적용할수있음이증명이되었다. 또한여러인종에서미토콘드리아 DNA 전체염기서열분석결과및각인종별유전적다양성에대한연구가진행되었고, 아프리칸미국인, 코카시안미국인, 히스패닉미국인, 에스토니안인종에서미토콘드리아 DNA 전체염기서열분석에따른유전적다양성을파악하기위한다양한연구들이진행되고있다. 그러나한국의경우, 법의학실무영역에서한국인에대한개인식별을위해서는한국인에서나타나는미토콘드리아 DNA의염기서열분석자료및유전적다양성에대한정보가필요하지만, 대부분미토콘드리아 DNA 조절부위또는조절부위의과변이부위, 암호부위의특정부위에국한되어있고, 전체염기서열에대한정보가매우제한적인상황이다. (Allard MW, et al., 2004, Jin HJ, et al, 2006, Jin HJ, et al., 2009, Lee HY, et al., 2006) 따라서한국인에서나타나는미토콘드리아 DNA 전체 36
48 염기서열에대한분석및그에따른유전적다양성에대한연구가 필요하다고보았다. 37
49 목적 1. 한국인에서차세대염기서열분서기법을이용하여미토콘드리아 DNA 전체염기서열을해독하고변이에대하여분석한다. 2. 한국인에서나타나는미토콘드리아 DNA 의변이의양상을 분석하고, 다른민족에대한연구를통해한국인의유전적다양성에 대하여분석한다. 3. 한국인의미토콘드리아 DNA 전체염기서열분석결과결정된 하플로그룹과하플로타입의양상을분석하고, 다른민족에대한연구를 통해한국인의유전적다양성에대하여분석한다. 4. 한국인의미토콘드리아 DNA 전체염기서열분석결과나타나는 점이형세포질성의양상을파악하고다른민족에대한연구를통해 한국의유전적다양성에대해분석한다. 5. 한국인에서미토콘드리아 DNA 의조절부위의염기서열분석결과와 전체염기서열분석결과를비교하여전체염기서열분석, 개세식별력등의 의의를알아본다. 38
50 재료와방법 1. 시료 한국인의미토콘드리아 DNA 조절부위 / 과변이부위에대한연구결과알려진모든하플로그룹을연구대상으로고려하기에는현실적으로제한이있었다. 따라서 6ㆍ25 한국전쟁과제주43사건관련유전자감식사업에서확보된것으로모두유전자검사에대한법적동의서가있는한국인시료중에서, 계통발생학적으로분류되는양상을고려하여, 이전연구보고자료와공용 mtdna 데이터베이스 (Jin HJ, et al., 2009, Lee HY, et al., 2006, 를바탕으로한국인에서흔하게나타나는것으로알려진하플로그룹에따라 186명의한국인시료를선별하였다. 하플로그룹결정은미토콘드리아 DNA 조절부위의과변위부위염기서열분석결과확인된변이의양상에근거한것이었다. 각시료는일부시료에서지역적및성별에대한정보를알수있었고, 선별된시료간의친족관계여부, 병력에대한정보는알수없었다. ( 표 1과표2 참조, 그림3과그림4, 그림5 참조 ). 매크로하플로그룹 D (macrohaplogroup D) 는한국인에서가장흔한하플로그룹으로, 미토콘드리아 DNA 조절부위염기서열에서확인되는변이양상만으로는하위하플로그룹을결정하기에는제한적이었다 (Allard MW, et al., 2004, Jin HJ, et al., 2009, Lee HY, et al., 2006), 이에따라하플로그룹 D4a, D4a3, D4b1, D5b (Haplogroup D4a, D4a3, D4b1, D5b) 를선별하였다. 하플로그룹 A, M7, F, G, B4, N9a 역시한국인에서흔히나타나는유형으로 (Allard MW, et al., 2004, Jin HJ, et al., 2009, Tanaka M, et al., 2004, Kivisild T, et al., 2002, Underhill PA, et al., 2007), 하플로그룹 A5a, M7a1, M7b1, F1b1, G1a1, B4a, B4c1c, N9a2a를선별하였다. 매크로하플로그룹 M9, G, D는미토콘드리아 DNA 조절부위의 16223, 16362, 489번뉴클레오티드위치에서공통적으로변이를가지고있는것이특징이기때문에조절부위에서확인되는변이양상만으로는하위하플로그룹을결정하기에제한적이고암호부위에서의정보가필요할것으로보아 (Jin HJ, et al., 2006, Lee HY, et al., 2006, Maruyama S, et al, 2013, 그림3), 이군에서선별하였다. 추가적으로제주도는한국본토와다른 39
51 유전적특성을가진것으로알려져있고 (Allard MW, et al., 2004), 하플로그룹 Y가 흔히 나타나는 것으로 알려져 있어서 세부 하플로그룹으로 분류가 가능한지 알아보기 위하여 이 군을 포함하여 선별하였다. 2. 미토콘드리아 DNA 추출과증폭 모든시료는냉동보관된혈액이었으며, 이들을해동한후 MaxwellR 16 Blood DNA Purification Kit (Promega, Madison, WI, USA) 와 MaxwellR 16 instrument (Promega) 을이용하여 DNA를추출하였다. DNA 증폭에사용할길잡이 (primer) 는미토콘드리아 DNA 전체를포함하는두개의앰플리콘 (amplicon) 이생성되도록구성하였으며, 길잡이의염기서열은기존에보고된연구결과 (Fendt L, et al., 2009, Gonder MK, et al., 2007) 를따랐다. ( 표 3 참조 ) 미토콘드리아 DNA를차세대염기서열분석법에적용하는과정에서이방법을사용할경우검출된염기서열의질과정확도를높이는것으로알려져있다. 각앰플리콘의중합효소연쇄반응에는 TaKaRa LA Taq DNA polymerase (Takara Bio Inc., Otsu, Shiga, Japan) 를사용하였으며, 각반응의결과물은 HiYieldTM Gel/PCR DNA mini Kit (Real Biotech Corporation, Taipei, Taiwan) 로정제하였다. 이들은 NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) 로정량한뒤각앰플리콘의농도가 50 ng/ μl이되도록희석하였으며, 각 20 μl씩같은양을혼합하여다음단계의실험에사용하였다. 모든실험장비와키트는각제조사에서제공한사용방법에따라적용하였다. 3. 라이브러리준비와초병렬염기서열분석 앞서준비된시료들에대하여, Ion XpressTM Plus Fragment Library Kit (Thermo Fisher Scientific) 를사용하여분절된라이브러리 (libaray) 를제작하였으며, DNA shearing은 kit에포함된 Ion Shear TM reaction을이용한효소에의한 DNA 분절화방법을 40
52 사용하였다. 각라이브러리에 Ion Xpress Barcode Adapters Kit (Thermo Fisher Scientific) 를이용해표지자를바코딩하였다. 이들을 Ion Library Quantitation Kit (Thermo Fisher Scientific) 을이용하여정량한뒤각각의농도가 26 pm이되도록희석하였다. 이들은미세구슬 (microbead) 에결합되어중합효소연쇄반응을거친뒤 PGM (personal genome machine) 에서염기서열분석이시행되었다. PGM은시료를주입한반도체칩을염기서열분석에사용하는데, 이실험에서는각각 16개의시료를올린 Ion 316 chip (Thermo Fisher Scientific) 을사용하였으며, 각시료세트에는대조검체인 SRM2392 (9947A) 이포함되어있었다. ( 그림6 참조 ) 모든장비와키트는제조사인 Thermo Fisher Scientific의가이드라인에따라사용되었다. 4. 자료분석 실험을통해얻은염기서열자료는 Ion Torrent Suite Software (Thermo Fisher Scientific) 를사용하여분석하였다. 기준염기서열로는 rcrs (the revised Cambridge Reference Sequence) 가사용되었으며, 이에 대한 변이 분석을 위하여 위 프로그램에서 제공하는 Variant Caller Plugin (Thermo Fisher Scientific) 이사용되었으며, 분석결과는 Variant Caller Format (VCF) 보고서로 제공되었다. 이와 더불어, Integrative Genomics Viewer (IGV) 를 사용하여 VCF 보고서에서 제외되었거나잘못검출된변이들을시각적으로재확인하였다. 하플로그룹은 PhyloTree Build 16 database (Behar DM, et al., 2012, van oven M, et al., 2009) 를 바탕으로, MitoTool ( 과 HaploGrep2 (Weissensteiner H, et al., 2016; 을이용하여결정하였다. 위에서얻은염기서열의변이와하플로그룹분석결과는 BAM 형태의 원자료를 mtdna-server (Weissensteiner H, et al., 2016; 를 통해 분석함으로써 다시 한 번 검증하였다. 5. 윤리적선언 41
53 본연구는서울대학교병원의생명연구원의의학연구윤리심의위원회에 의해승인을받았으며, 사전동의를받았음을확인받았다. 또한모든 시료는유전자검사에대하여법적동의서를받은것이었다. 42
54 연구결과 1. 미토콘드리아 DNA 염기서열분석에서 coverage 1.1 염기서열분석과 coverage 염기서열분석에서 coverage란해당 DNA 염기서열을몇번해독했는지를평균적인수치로표현하는것으로, 알고있는표준염기서열에맞추어정렬한대로해독한다는의미로서 cover 에서유래하였다. 차세대염기서열분석기법의특성상특정염기서열위치에서특정변이가확인되었다는신뢰할만한염기검출 (base call) 결과를얻기위해서는여러번의해독이필요하고, 해독이모든염기서열부위에서동일한횟수로일어나는것이아니라부위에따라달라질수있기때문에이를확인하는것은중요하다. 1.2 염기서열부위와 coverage 본연구에서선별된모든 186명의시료에서미토콘드리아 DNA 전체염기서열의 coverage는평균적으로 1,200배였다. 선별된모든개체의미토콘드리아 DNA를해독하였을때, 2,600번째뉴클레오티드가위치한부위부근의크기 300 염기쌍이하의염기서열부위에서 8,000배에이르는유달리높은 coverage를보여주었다. 대조적으로 13,600번째뉴클레오티드와 14,800번째뉴클레오티드사이에존재하는염기서열부위에서는 500배이하의낮은 coverage를보였다. 2. 미토콘드리아 DNA 전체염기서열에서변이 2.1 변이검출및불일치해석 차세대염기서열분석기법을적용하여얻은분석결과를그래프뷰어를 사용하여확인하였다. 이는차세대염기서열분석결과에대하여이해하고 43
55 염기서열분석중잡음을구분할뿐만아니라변이검출의불일치를해석하는데유용하였다. (Parson W, et al., 2013) 변이검출형식보고서 (Variant caller format; VCF) 에서보고된변이는 Integrative Genomics Viewer (IGV) 와 mtdna-server를통해전체염기서열을수작업으로일일이다시확인하였다. (Weissensteiner H, et al., 2016) 이를통해 IGV에서검출되지않았거나오검출된변이의유무를확인할수있었다. ( 표 4, 표 5 참조 ) 2.2 보고되지않은변이오류 VCF 에보고되지는않았으나기존에가지고있었던생거방식을통한 조절부위염기서열분석자료로변이의유무를확인할수있었으며그 결과는다음과같았다. 71번째뉴클레오티드위치변이로검출되지않았으나, 실제염기서열에서는이위치에뉴클레오티드 ( 구아닌 ) 가추가로삽입된변이 (71.1G) 가확인되었다. 하플로그룹 B4a군 (B4a1c1a, B4a1c1a1) 에해당하는 3명의개체에서이변이가확인되었다. 455번째뉴클레오티드위치 변이로 검출되지 않았으나, 실제 염기서열에서는 이 위치에 뉴클레오티드 ( 티민 ) 가 추가로 삽입된 변이 (455.1T) 가 확인되었다. 하플로그룹 D4(D4a, D4d) 에 해당하는 3명의 개체에서 이 변이가 확인되었다. 573번째뉴클레오티드위치변이로검출되지않았으나, 실제염기서열에서는이위치에뉴클레오티드 ( 시토신 ) 가추가로삽입된변이 (573.1C) 가확인되었다. 하플로그룹 A5a(A5a3) 에해당하는 1명의개체, 하플로그룹 M9;G;D군 (D4e2a) 에해당하는 2명의개체에서이변이가확인되었다. IGV 에서확인된변이역시 VCF 에보고되지않아서위음성오류로 확인된변이는다음과같았다. 44
56 960번째뉴클레오티드위치 변이로 검출되지 않았으나, 실제 염기서열에서는 이 위치에 뉴클레오티드 ( 시토신 ) 가 추가로 삽입된 변이 (960.1C) 가 확인되었다. 하플로그룹 N(N9a2a) 에 해당하는 11명의 개체에서 이 변이가 확인되었다 번째와 16183번째뉴클레오티드위치 : 변이로검출되지않았으나, 실제염기서열에서는이위치에뉴클레오티드 ( 아데노신 시토신 ) 가치환된변이 (16182C, 16183C) 가확인되었다. 하플로그룹 B4군에해당하는전체 20명의개채, 하플로그룹 F1b1군에해당하는전체 16명의개체, 하플로그룹 M7b군 (M7b1a1a1) 에해당하는 1명의개체에서이변이가확인되었다. 하플로그룹 M7b군 (M7b1a1a1) 에해당하는다른 1명의개체, 하플로그룹 D군 (D5b) 에해당하는 2명의개체에서 16183번의변이만관찰되었다. 2.3 삽입 - 결실과관련된정렬오류 삽입 - 결실과관련된정렬오류 (Indel-related alignment errors) 에서도 VCF 에서보여진결과와실제염기서열을확인하였을 때의결과에다음과같은차이가있었다. 249번째뉴클레오티드위치의결실변이 247번째뉴클레오티드위치에결실이있는변이로검출 (247GA 247G) 되었으나, 실제염기서열을확인하였을때 249번째뉴클레오티드 ( 아데노신 ) 가결실된변이 (249del) 였다. 이변이는하플로그룹 F1b1군전체 16명의개체에서확인되었다 번째뉴클레오티드위치의결실변이 513번째뉴클레오티드와이웃한뉴클레오티드 2곳에서결실이있는변이로검출 (513GCA 513G) 되었으나, 실제염기서열을확인하였을때 523번째와 524번째뉴클레오티드 ( 각각아데노신, 시토신 ) 가결실된변이 ( del) 였다. 이변이는하플로그룹 A5a 전체 20명의개체, 하플로그룹 B4a 전체 16명의개체, 하플로그룹 D4b와 M9;G;D(D4b1, D4b2a2, D4d) 에해당하는 11명의개체, 하플로그룹 F1b1 전체 45
57 16 명의개체, 하플로그룹 G1a1 에해당하는 1 명의개체, 하플로그룹 M7a1 전체 9 명의개체에서확인되었다. 2,156번째뉴클레오티드위치삽입변이 2,150번째뉴클레오티드에추가로뉴클레오티드 ( 아데노신 ) 가삽입된변이로검출 (2150T 2150TA) 되었으나, 실제염기서열을확인하였을때 2,159번째에추가로뉴클레오티드 ( 아데노신 ) 가삽입된변이 (2156.1A) 였다. 이변이는하플로그룹 A5a 전체 20명의개체에서확인되었다. 5,899번째뉴클레오티드위치삽입변이 5,894번째뉴클레오티드위치에서삽입된변이 (5894A 5894AC) 로검출되었으나, 실제염기서열을확인하였을때 5,899번째에뉴클레오티드 ( 시토신 ) 가삽입된변이 (5899.1C) 였다. 이변이는하플로그룹 M7a1 전체 9명의개체에서확인되었다. 8,281-8,289번째뉴클레오티드위치결실변이 8,270번째뉴클레오티드위치에서여러개의뉴클레오티드가결실된변이 (8270CACCCCCTCT 8270C) 로검출되었으나, 실제염기서열을확인하였을때 8,281번째에서 8,289번째사이에위치하는뉴클레오티드 (CCCCCTCTA) 가결실된변이 ( del) 로확인되었다. 이변이는하플로그룹 B군 (B4a, B4c) 전체 20명의개체에서확인되었다. poly-tracts나삽입-결실로인한오정렬에따른혼합결과는 mtdna server를통해확인하였었다. 이러한불일치는이위치부근에 poly C tract나 poly A tract으로인한정렬오류에의한것이었다. (Zhou Y, et al., 2016) 4,248번째뉴클레오티드위치치환변이 4,246번째뉴클레오티드부근의염기서열이혼합결과 (4246ATT AT,ATC) 로검출되었으나, 실제염기서열을확인하였을때 4,248번째에뉴클레오티드 ( 아데노신 시토신 ) 가치환된변이 (4248C) 였다. 이변이는하플로그룹 A5a 전체 20명의개체에서확인되었다. 46
58 9,540번째뉴클레오티드위치변이 9,540번째뉴클레오티드부근의염기서열이혼합결과 (9540C 또는 9540T/C) 로검출되었으나, 실제염기서열을확인하였을때 9,540번째뉴클레오티드 ( 티민 시토신 ) 으로치환된변이 (9540C) 였다. 이변이는하플로그룹 D4군에해당되는 11명의개체에서확인되었다. 2.4 기타변이 5명의시료에서확인된 2,478번뉴클레오티드위치, 20명의시료에서확인된 10,838번뉴클레오티드위치는 IGV를사용하였을때혼합부위로관찰되었으나, mtdna-server에서는그렇지않았다. 이러한소견들은특이하플로그룹에서이들부위부근에 polymeric tracts가결여되어생긴인공산물때문인것으로추정되었다. 9,824번뉴클레오티드위치에서 3개의대립형질 (tri-allele) 변이가확인되었다. 이들은하플로그룹 M7에서시토신전이 (transition), 하플로그룹 M9;G;D에서아데노신전이 (transition) 였다. 2.5 동형세포질성변이 186명의한국인시료에서총 6,495 변이가관찰되었다. 이중 159개의변이는삽입결실 (2.45%) 이고나머지 6,336개의변이 (97.55%) 는치환에의한변이였다. 미토콘드리아 DNA 전체염기서열에서확인되는동형세포질성변이들을조절부위와암호부위에따라나타나는변이의양상을분석해보았다. 동형세포질성변이들은암호부위에서더높은빈도로발견되었으며, 평균적으로암호부위에서 74.10%, 조절부위에서 25.90% 였다. ( 표 6 참조 ) 하플로그룹 D4 모두 1,998개의변이가관찰되었으며, 조절부위에서 446개 (22.32%), 암호부위에서 1,552개 (77.68%) 로, 조절부위의변이의개수에대하여암호부위의변이의개수의비율은 3.48로나타났다. 하플로그룹 A5a 47
59 모두 611 개의변이가관찰되었으며, 조절부위에서 168 개 (27.50%), 암호부위에서 443 개 (72.50%) 로, 조절부위의변이의개수에대하여 암호부위의변이의개수의비율은 2.64 로나타났다. 하플로그룹 M7 (M7a1, M7b1) 모두 774개의변이가관찰되었으며, 조절부위에서 168개 (21.71%), 암호부위에서 606개 (78.29%) 로, 조절부위의변이의개수에대하여암호부위의변이의개수의비율은 3.61로나타났다. 하플로그룹 F1b1 모두 609개의변이가관찰되었으며, 조절부위에서 212개 (34.81%), 암호부위에서 397개 (65.19%) 로, 조절부위의변이의개수에대하여암호부위의변이의개수의비율은 1.87로나타났다. 하플로그룹 G1a1 모두 765개의변이가관찰되었으며, 조절부위에서 171개 (22.35%), 암호부위에서 594개 (77.65%) 로, 조절부위의변이의개수에대하여암호부위의변이의개수의비율은 3.47로나타났다. 하플로그룹 B4 (B4a, B4c1c) 모두 513개의변이가관찰되었으며, 조절부위에서 204개 (39.77%), 암호부위에서 309개 (60.23%) 로, 조절부위의변이의개수에대하여암호부위의변이의개수의비율은 1.51로나타났다. 하플로그룹 N9a2a 모두 299개의변이가관찰되었으며, 조절부위에서 97개 (32.44%), 암호부위에서 202개 (67.56%) 로, 조절부위의변이의개수에대하여암호부위의변이의개수의비율은 2.08로나타났다. 하플로그룹 M9;G;D 모두 565개의변이가관찰되었으며, 조절부위에서 101개 (17.88%), 암호부위에서 464개 (82.12%) 로, 조절부위의변이의개수에대하여암호부위의변이의개수의비율은 4.59로나타났다. 하플로그룹 Y1 군 48
60 모두 361 개의변이가관찰되었으며, 조절부위에서 115 개 (31.86%), 암호부위에서 246 개 (68.14%) 로, 조절부위의변이의개수에대하여 암호부위의변이의개수의비율은 2.14 로나타났다. 각하플로그룹에서의변이의비율은다양해서하플로그룹 B4의경우 가장 작은 비율 (1.51) 을 보여주었으며 ( 암호부위에서 60.23%, 조절부위에서 39.77%), 하플로그룹 M9;G;D에서 가장 높은 비율 (4.59) 을 보여주었다 ( 암호부위에서 82.12%, 조절부위에서 17.88%). 2.6 공통변이 186 명의한국인각개체에서미토콘드리아 DNA 전체염기서열 분석결과확인된변이중각개체에서전체또는대부분에서공통적으로 관찰되는변이가확인되었다. 모든개체에서공통적으로관찰된변이 73번, 263번, 750번, 1438번, 4769번, 7028번, 11719번, 14766번, 15326번째뉴클레오티드위치에관찰되었다. (73G, 263G, 750G, 1438G, 4769G, 7028G, 11719A, 14766T, 15326G) 약절반이상의개체에서공통적으로관찰된변이 489번, 2706번, 8701번, 9540번, 10398번, 10400번, 10873번, 12705번, 14783번, 15043번, 15301번, 16223번, 16519번째뉴클레오티드위치에서발견되었다. (489C, 2706G, 8701G, 9540C, 10398G, 10400T, 10873C, 12705T, 14783C, 15043A, 15301A, 16223T, 16519C) 2.7 개별변이 (private mutation) 모든한국인개체의미토콘드리아 DNA 전체염기서열분석결과같은하플로그룹안에서도다르게나타나는개별변이가관찰되었다. HaploGrep을이용한분석결과세부하플로그룹결정에관여되는변이는아니지만지역적또는세계적인개체변이로간주되는변이들이 49
61 있었으며, 이들중 2 개체이상에서공통적으로나타나는변이들을 관찰할수있었다. 각하플로그룹안에서공통적으로관찰된개별변이는 다음과같다. ( 표 7 참조 ) 하플로그룹 D4a1 14,926 번째뉴클레오티드위치에서개별변이 (14926G) 가전체 11 개체중 2 개체에서관찰되었다. 하플로그룹 D4b1a1 4,113번째 뉴클레오티드 위치에서 개별변이 (4113A), 4,646번째 뉴클레오티드 위치에서 개별변이 (4646C) 가 전체 6개체에서 모두 관찰되었다. 하플로그룹 D4d 6,524 번째뉴클레오티드위치에서개별변이 (6524C) 가전체 4 개체 중 2 개체에서관찰되었다. 하플로그룹 A5a 5,250 번째뉴클레오티드위치에서개별변이 (5250C) 가전체 6 개체 중 2 개체에서관찰되었다. 하플로그룹 A5a1a 8,619 번째뉴클레오티드위치에서개별변이 (8619T) 가전체 11 개체 중 2 개체에서관찰되었다. 하플로그룹 F1b1a2 3,395 번째뉴클레오티드위치에서개별변이 (3395G) 가전체 4 개체 중 2 개체에서관찰되었다. 하플로그룹 G1a1a 9,254번째뉴클레오티드위치에서개별변이 (9254G) 가전체 11개체중 2개체에서관찰되었다. 9,410번째뉴클레오티드위치에서개별변이 (9410G) 가전체 11개체중 2개체에서관찰되었다. 9,494번째뉴클레오티드위치에서개별변이 (9494G) 가전체 11개체중 2개체에서관찰되었다. 50
62 하플로그룹 G1a1a1 1,822번째뉴클레오티드위치에서개별변이 (1822C), 9,854번째뉴클레오티드위치에서개별변이 (9854C) 가전체 8개체중 6개체에서관찰되었다. 하플로그룹 B4a 15,926 번째뉴클레오티드위치에서개별변이 (15926T) 가전체 4 개체 중 4 개체에서관찰되었다. 하플로그룹 B ,863 번째뉴클레오티드위치에서개별변이 (6863G) 가전체 2 개체중 2 개체에서관찰되었다. 하플로그룹 B4a1c1a 8,595 번째뉴클레오티드위치에서개별변이 (8595T) 가전체 4 개체중 2 개체에서관찰되었다. 하플로그룹 N9a2a 3,866 번째뉴클레오티드위치에서개별변이 (3866C) 가전체 10 개체 중 3 개체에서관찰되었다. 하플로그룹 Y1 9,254 번째뉴클레오티드위치에서개별변이 (9254G) 가전체 9 개체 중 2 개체에서관찰되었다. 3. 하플로그룹분류 14개 (D4a, D4a3, D4b1, D5b, A5a, M7a1, M7b1, F1b1, G1a1, B4a, B4c1c, N9a2a, M9;G;D, Y1) 의하플로그룹 ( 조절부위근거 ) 에해당되었던 186 개체의각미토콘드리아 DNA 전체염기서열분석결과확인된변이는 HaploGrep를이용하여 Phylotree 16에따라하위하플로그룹으로분류할수있었다. ( 표 8 참조 ) 계통발생학나무를따라 51
63 각하플로그룹을결정하는주요변이는표 9 에제시하였다 하플로그룹 D (D4a, D4a3, D4b1, D5b) 하플로그룹 D4a(40개체 ) 는미토콘드리아 DNA 전체염기서열분석을통해 D4a(3개체 ), D4a1(11개체 ), D4a1a1(2개체 ), D4a1b(10개체 ), D4a1c(4개체 ), D4a2(8개체 ), D4a2a(1개체 ), D4f1(1개체 ) 등모두 8유형의하위하플로그룹으로세분화할수있었다. 하플로그룹 D4a3, D4b1a1, D5b의경우미토콘드리아 DNA 전체염기서열분석에도불구하고추가적으로세분화된하위하플로그룹은없었으나, 하위하플로그룹인 D4a3b, D4b1a1, D5b1b 등으로각각세분화될수있었다. 각개체별염기서열및변이를분석하여보았을때, 같은하플로그룹내에서도서로다른변이가관찰되었다. 하플로그룹 D4a1 11개체중 3개체와 2개체에서각각서로동일한염기서열및변이를보여주었고나머지 6개체에서는서로다른일부변이가관찰되었다. 하플로그룹 D4a1b 역시 10개체중 3개체와 2개체에서각각서로동일한염기서열및변이를보여주었고나머지 4개체에서는서로다른일부변이가관찰되었다. 하플로그룹 D4a2 8개체중 4개체와 2개체에서각각서로동일한염기서열및변이를보여주었고나머지 2개체에서는서로다른일부변이가관찰되었다. 하플로그룹 D4b1 6개체중 3개체에서서로동일한염기서열및변이를보여주었고나머지 3개체에서는서로다른일부변이가관찰되었다. 이외하플로그룹의각개체는서로다른일부변이가관찰되었다 하플로그룹 A5a 하플로그룹 A5a(20개체 ) 는전체염기서열분석에서확인된변이를통해하플로그룹 A5a(6개체 ), A5a1a(11개체 ), A5a1a1(1개체 ), A5a1a2(1개체 ), A5a3(1개체 ) 등의하위하플로그룹으로세분화할수있었다. 하플로그룹 A5a의각개체의염기서열을비교분석하였을때, 6개체중 2개체와 2개체에서각각서로동일한염기서열및변이를보여주었고나머지 2개체에서는서로다른일부변이가관찰되었다. 52
64 하플로그룹 A5a1a 11개체중 2개체와 2개체에서각각서로동일한염기서열및변이를보여주었고나머지 7개체에서는서로다른일부변이가관찰되었다. 이외하플로그룹의각개체는서로다른일부변이가관찰되었다. 3.3 하플로그룹 M7 군 (M7a1, M7b1) 하플로그룹 M7a1(9개체 ) 는전체염기서열분석결과 M7a1a(4개체 ), M7a1a1(1개체 ), M7a1a5a(1개체 ), M7a1a9(3개체 ) 등의하위하플로그룹으로세분화할수있었다. 하플로그룹 M7b1(10개체 ) 은대부분이하위하플로그룹 M7b1a1a1 (9개체) 으로세분화되었고, 1개체에서하위하플로그룹 M7b1a1a로세분화될수있었다. 각하플로그룹의각개체의염기서열을분석하였을때, 하플로그룹 M7a1a 4개체중 2개체는서로동일한염기서열및변이를보여주었고나머지 2개체에서는서로다른일부변이가관찰되었다. 이외하플로그룹의각개체는서로다른일부변이가관찰되었다 하플로그룹 F1b1 하플로그룹 F1b1(16개체 ) 는 전체염기서열분석결과 하플로그룹 F1b1a(1개체 ), F1b1a1(4개체 ), F1b1a1a(4개체 ), F1b1a1a1(1개체 ), F1b1a1a2(2개체 ), F1b1a2(4개체 ) 등의 하위 하플로그룹으로 세분화되었다. 각하플로그룹의각개체의염기서열을분석하였을때, 하플로그룹 F1b1a1 4개체중 2개체는서로동일한염기서열및변이를보여주었고 나머지 2개체에서는 서로 다른 일부 변이가 관찰되었다. 하플로그룹 F1b1a1a 4개체 중 3개체는 서로 동일한 염기서열 및 변이를 보여주었다. 이외 하플로그룹의 각 개체는 서로 다른 일부 변이가 관찰되었다 하플로그룹 N9a2a 하플로그룹 N9a2a(11 개체 ) 는전체염기서열분석결과에도하플로그룹 53
65 분류는대부분변화가없었고 (10개체), 1개체에서하위하플로그룹 N9a2a2로세분화될수있었다. 각하플로그룹의각개체의염기서열을분석하였을때, 하플로그룹 N9a2a 11개체중 2개체, 2개체는각각서로동일한염기서열및변이를보여주었고, 나머지 7개체에서는서로다른일부변이가관찰되었다 하플로그룹 G1a1 하플로그룹 G1a1(19개체 ) 은전체염기서열분석결과하위하플로그룹 G1a1a(11개체 ), G1a1a1(8개체 ) 로세분화할수있었다. 4793G, 11914A, 827G 변이는하플로그룹 G1a1a와 G1a1a1을구분하게하는중요한변이였다. 각하플로그룹의각개체의염기서열을분석하였을때, 하플로그룹 G1a1a 11개체중 2개체는서로동일한염기서열및변이를보여주었고나머지 9개체에서는서로다른일부변이가관찰되었다. 하플로그룹 G1a1a1 8개체중 3개체는서로동일한염기서열및변이를보여주었고나머지 5개체에서는서로다른일부변이가관찰되었다. 이외하플로그룹의각개체는서로다른일부변이가관찰되었다 하플로그룹 Y1 하플로그룹 Y1(12개체 ) 은대부분 Y1(9개체 ) 로분류되었고, 이외하플로그룹 Y1a(1개체 ), Y1b1(1개체 ), Y1b1a(1개체 ) 로세분화될수있었다. 각하플로그룹의각개체의염기서열을분석하였을때, 하플로그룹 Y1 9개체중 2개체는서로동일한염기서열및변이를보여주었고나머지 7개체에서는서로다른일부변이가관찰되었다 하플로그룹 B4 (B4a, B4c1c) 하플로그룹 B4a(16 개체 ) 는전체연기서열분석결과 B4a(4 개체 ), B (2 개체 ), B4a1c1a(4 개체 ), B4ac1a1(6 개체 ) 로분류할수 있었다. 54
66 하플로그룹 B4a군에서미토콘드리아 DNA 조절부위에서확인되는변이만으로는하위하플로그룹분류가제한적이었으며, 전체염기서열에서확인되는변이양상을통해하위하플로그룹유형결정이가능하였다. MitoTool을이용하여조절부위변이를근거로분석하였을때, 하플로그룹 B4a의 16개체중 13개체가여러후보하위하플로그룹으로제시되었는데 (B4a; B4a1c1a1; B4i or B4a; B4i), 하위하플로그룹으로분류해주는기준이되는일부변이가관찰되지않음으로계통발생학적나무에서상위즉조상의계열에가까운것으로간주되었기때문으로생각되었다. (Zhou Y, et al, 2016) 미토콘드리아 DNA 전체염기서열에서나타나는변이양상을분석하였을때, 9개체가하나의하위하플로그룹 (B4a; B4a1c1a1; B4i -> B4a1c1a1, B4a;B4i -> B4a) 으로분류될수있었다. HaploGrep을이용하여분석한경우, 조절부위에서확인되는변이로하플로그룹을분류한경우, 10개체가하플로그룹 B 이었으나, 전체염기서열에서확인된변이로분석하였을때, 4개의하위하플로그룹 (B4a, B , B4a1c1a, B4a1c1a1) 으로분류될수있었다. 하플로그룹 B4a의모든개체에서조절부위에서확인되는변이만근거로분석하였을때와전체염기서열에서확인되는변이를근거로분석하였을때, 각각전반적인질점수 (quality score) 는 0.874에서 0.943으로증가하였다. 각하플로그룹의각개체의염기서열을분석하였을때, 하플로그룹 B4a군 4개체에서 2개체는서로동일한염기서열및변이가관찰되었고나머지 2개체는서로다른일부변이가관찰되었다. 하플로그룹 B 의경우 2개체모두동일한염기서열및변이가관찰되었다. 이외하플로그룹의각개체는서로다른일부변이가관찰되었다 하플로그룹 M9;G;D 하플로그룹 M9;G;D로분류되었던 17개체는전체염기서열분석결과 D4b2a2(1개체 ), D4d(4개체 ), D4e2(2개체 ), D4e2a(2개체 ), D4f1(3개체 ), D4j(5개체 ) 로재분류할수있었다. 매크로하플로그룹 M9, G, D는조절부위에서확인되는변이는하위하플로그룹분류에는거의유용하지않았는데하위하플로그룹분류에중요한기준변이중조절부위에서확인되는변이는적기때문이다. (Jin HJ, et al., 2006, Lee HY, et al., 2006) 17개체에서미토콘드리아 DNA 55
67 전체염기서열에서확인된변이가있어야하위하플로그룹분류가가능하였다 (D4b2a2, D4d, D4e2, D4e2a, D4f1, D4j). 전체염기서열분석을통해암호부위에서확인되는변이의정보는매크로하플로그룹 M9, G, D의하위하플로그룹분류에있어서중요할것으로생각된다. 각하플로그룹의각개체의염기서열을분석하였을때, 각세부하플로그룹으로분류된각개체는서로다른일부변이가관찰되었다. 4. 하플로타입 (Haplotype) 186명의한국인의미토콘드리아 DNA의조절부위와전체염기서열분석결과각각나타나는하플로타입을분석하였다. 각염기서열을비교하였을때 2개이상의변이가차이가날때고유의하플로타입으로정의하였다. (Nilson M, et al., 2008, Parson W, et al., 2014) 조절부위에근거하였을때 70개의하플로타입은전체염기서열분석결과 137개의하플로타입으로나타났다. ( 표 10 참조 ) 하플로그룹 D4a 40 개체에서조절부위로만했을때하플로타입은 11 개였으나 전체염기서열로하였을때 26 개의하플로타입으로나타났다. 하플로그룹 D4a3 3 개체에서조절부위로만하였을때, 하플로타입은 1 개였으나전체 염기서열로하였을때 3 개의하플로타입으로나타났다. 하플로그룹 D4b1 6 개체에서조절부위로만하였을때, 하플로타입은 1 개였으나전체 염기서열로하였을때 2 개의하플로타입으로나타났다. 하플로그룹 D5b 3 개체에서조절부위로만하였을때, 하플로타입은 2 개였으나전체 염기서열로하였을때 3 개의하플로타입으로나타났다. 56
68 하플로그룹 A5a 20 개체에서조절부위로만하였을때, 하플로타입은 12 개였으나 전체염기서열로하였을때 14 개의하플로타입으로나타났다. 하플로그룹 M7a1 9 개체에서조절부위로만하였을때, 하플로타입은 2 개였으나전체 염기서열로하였을때 8 개의하플로타입으로나타났다. 하플로그룹 M7b1 10 개체에서조절부위로만하였을때, 하플로타입은 6 개였으나전체 염기서열로하였을때 10 개의하플로타입으로나타났다. 하플로그룹 F1b1 16 개체에서조절부위로만하였을때, 하플로타입은 2 개였으나전체 염기서열로하였을때 11 개의하플로타입으로나타났다. 하플로그룹 G1a1 19 개체에서조절부위로만하였을때, 하플로타입은 8 개였으나전체 염기서열로하였을때 15 개의하플로타입으로나타났다. 하플로그룹 B4a 16 개체에서조절부위로만하였을때, 하플로타입은 8 개였으나전체 염기서열로하였을때 11 개의하플로타입으로나타났다. 하플로그룹 B4c1c 4 개체에서조절부위, 전체염기서열분석결과모두동일하게 1 개의 하플로타입으로나타났다. 하플로그룹 N9a2a 11 개체에서조절부위로만하였을때, 하플로타입은 3 개였으나전체 염기서열로하였을때 9 개의하플로타입으로나타났다. 하플로그룹 M9;G;D 17 개체에서조절부위로만하였을때, 하플로타입은 7 개였으나전체 57
69 염기서열로하였을때 15 개의하플로타입으로나타났다. 하플로그룹 Y1 12 개체에서조절부위로만하였을때, 하플로타입은 6 개였으나전체 염기서열로하였을때 9 개의하플로타입으로나타났다. 5. 점이형세포질성 (Point heteroplasmy) 5.1. 점이형세포질성의정의 점이형세포질성은다음과같은기준을만족하는경우로정의하였다. 1) 이형세포질성을이루는변이들중부요소 (minor component) 가적어도 40배이상의 coverage를가져야하며 (Jin HJ, et al., 2006), 2) 미토콘드리아 DNA와유사한염기서열을갖는핵 DNA (nuclear mitochondrial DNA, NUMT) 가함께증폭된것으로의심되는경우는제외하며, 3) mtdna server의분석결과해당이형세포질성이신뢰할만한결과로확인된것이었다 (Weissensteiner H, et al., 2016) 점이형세포질성에서 coverage 이형세포성을구성하는부요소 (minor component) 중최소값은하플로그룹 F1b1a1a에속하는한개체의 2561번째뉴클레오티드로, 그비율은 0.61%, coverage는 40배였으며, 최대값은하플로그룹 A5a에속하는한개체의 13704번째뉴클레오티드로비율은 45.93%, coverage는 95배였다. ( 표 11 참조 ) 5.3. 점이형세포질성의분포와양상 한국인 186개체의시료에서점이형세포질성에대한분석결과, 이중 40개체 (21.51%) 에서 52곳의이형세포질성이관찰되었다. 부위별로는 조절부위에서 16개 (30.77%), 암호부위에서 36개 (69.23%) 가 관찰되었다 ( 표 11, 표 12 참조 ). 7개체에서 2개 이상의 점 이형세포질성을 가진 것으로 나타났으며, 1개체의 경우 5개의 점 58
70 이형세포질성이있었던것으로확인되었다. 염기서열위치로보았을때, 64번, 2561번뉴클레오티드위치에서 2번반복해서관찰되었고, 12개의이형세포질성변이가 16039번에서 16234번사이에몰려서위치하고나머지 40개가 64번에서 15987사이에존재하는고르지않은분포를보였다. 대개조절부위에비해암호부위에서이형세포질성이 3-4배이상의빈도로발생함을관찰하였으나, 하플로그룹 G1a1과 B4에서는양쪽부위에서유사한빈도로발생하였음을관찰하였다 하플로그룹 D4 모두 11 개체에서 13 곳의이형세포질성이관찰되었다. 하플로그룹 D4a 1개체에서 3곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치로는 2844G/A( 주요소 / 부요소, 2.18%), 5973G/A (2.76%), 16126T/C (4.82%) 등이었다. 하플로그룹 D4a1 3개체에서각각 1곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치로는 9457T/C(7.81%), 9742C/T(9.87%), 16103A/G(5.68%) 등이었다. 하플로그룹 D4a1b 2개체에서 각각 1곳의 이형세포질성 변이가 관찰되었으며, 뉴클레오티드 위치로는 990T/C(3.12%), 2552T/C(1.12%) 등이었다. 하플로그룹 D4a1c 2개체에서 각각 1곳의 이형세포질성 변이가 관찰되었으며 뉴클레오티드 위치로는 114C/T(5.11%), 4088C/T(27.93%) 등이었다. 하플로그룹 D4a2a 1개체에서 1곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치로는 9064G/A(11.54%) 등이었다. 59
71 하플로그룹 D4a3b 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 6325C/T(37.69%) 등이었다. 하플로그룹 D4b1a1 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치로는 64C/T(26.04%) 등이었다 하플로그룹 A5a 모두 4 개체에서 4 곳의이형세포질성이관찰되었다. 하플로그룹 A5 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 13704T/C(45.93%) 등이었다. 하플로그룹 A5a1a 3개체에서각 1곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 2383T/C(12.8%), 4050T/C(16.03%), 16094C/T(2.79%) 등이었다 하플로그룹 M7 모두 5 개체에서 5 곳의이형세포질성이관찰되었다. 하플로그룹 M7a1a 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 13810A/G(22.37%) 등이었다. 하플로그룹 M7a1a5a 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 1425T/C(2.45%) 등이었다. 60
72 하플로그룹 M7a1a9 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 4728A/G(10.49%) 등이었다. 하플로그룹 M7b1a1a 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 64C/T(17.66%) 등이었다. 하플로그룹 M7b1a1a1 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 15686A/G(23.93%) 등이었다 하플로그룹 F1b1 하플로그룹 F1b1a1a 의 1 개체에서 3 곳의이형세포질성변이가 관찰되었으며, 뉴클레오티드위치는 2561T/C(0.61%), 2644A/G(1.02%), 10503T/C(6.05%) 등이었다 하플로그룹 G1a1 모두 4 개체에서 9 곳의이형세포질성변이가관찰되었다. 하플로그룹 G1a1a 1 개체에서 2 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 2580T/C(2.77%), 7606A/G(7.3%) 등이었다. 하플로그룹 G1a1a 1개체에서 5곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는, 12361A/G(18.75%), 16111C/T(17.68%), 16140T/C(18.24%), 16223T/C(21.67%), 16234C/T(19.88%) 등이었다. 하플로그룹 G1a1a 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 61
73 뉴클레오티드위치는 16129A/G(12.35%) 등이었다. 하플로그룹 G1a1a1 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 15987C/T(18.36%) 등이었다 하플로그룹 B4 모두 6 개체에서 6 곳의이형세포질성변이가관찰되었다. 하플로그룹 B4a 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 9618C/A(2.73%) 등이었다. 하플로그룹 B 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 6892G/A(2.7%) 등이었다. 하플로그룹 B4a1c1a 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 8805G/A(29.28%) 등이었다. 하플로그룹 B4a1c1a1 3개체에서각각 1곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 93A/G(3.79%), 16092T/C(3.1%), 16150C/T(9.58%) 등이었다 하플로그룹 N9a2a 모두 4 개체에서 6 곳의이형세포질성변이가관찰되었다. 하플로그룹 N9a2a 1 개체에서 2 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 9137T/C(3.66%), 11884A/G(4.32%) 62
74 등이었다. 하플로그룹 N9a2a 2개체에서각각 1곳의이형세포질성변이가관찰되었으며, 각각 뉴클레오티드 위치는 3335T/C(11.04%), 12868G/A(2.21%) 등이었다. 하플로그룹 N9a2a2 1개체에서 2곳의 이형세포질성 변이가 관찰되었으며, 뉴클레오티드 위치는 896A/G(4.98%), 16129G/A(7.23%) 등이었다 하플로그룹 M9;G;D 모두 5 개체에서 6 곳의이형세포질성변이가관찰되었다. 하플로그룹 D4e2a 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 2561T/C(0.78%) 등이었다. 하플로그룹 D4f1 1 개체에서 1 곳의이형세포질성변이가관찰되었으며, 뉴클레오티드위치는 6261G/A(4.74%) 등이었다. 하플로그룹 D4j 1개체에서 2곳의 이형세포질성 변이가 관찰되었으며, 뉴클레오티드 위치는 3062T/C(17.53%), 4710T/C(8.3%) 등이었다. 하플로그룹 D4j 2개체에서 각각 1곳의 이형세포질성 변이가 관찰되었으며, 뉴클레오티드 위치는 11598C/T(11.66%), 16039G/A(8.94%) 등이었다. 63
75 하플로그룹 Y1 하플로그룹 Y1 에서는이형세포질성변이가관찰되지않았다 동일개체에서나타나는이형세포질성변이 동일개체에서나타나는이형세포질성의뉴클레오티드구성을분석해보았다. 3개의이형세포질성변이가있던 2개의개체에서각각 T/C (x2) A/G, G/A (x2) T/C로, 5개의이형세포질성변이가있던개체에서는 T/C (x2) C/T (x2) A/G로관찰되었다. 2개의이형세포질성변이가있던 4개체에서는각각 A/G G/A, T/C (x2), T/C (x2), T/C A/G로구성되어있었다 미토콘드리아염기서열부위와이형세포질성변이 조절부위에서는과변이부위 I에 12개의과변이부위 II에 4개의이형세포질성변이가관찰되었다. 암호부위에서는 12S rrna 유전자부위에서 3개, 16S rrna 유전자부위에서 8개, NADH dehydrogenase 1 유전자부위에서 3개, NADH dehydrogenase 2 유전자부위에서 2개, Cytochrome c oxidase 1 유전자부위에서 4개, Cytochrome c oxidase 2 유전자부위에서 1개, ATP synthase 6 유전자부위에서 3개, Cytochrome c oxidase 3 유전자부위에서 3개, NADH dehydrogenase 4L 유전자부위에서 1개, NADH dehydrogenase 4 유전자부위에서 2개, NADH dehydrogenase 5 유전자부위에서 4개, Cytochrome B 유전자부위에서 1개 trna Pro에서 1개가관찰되었다 하플로그룹을결정하는변이부위에형성된 점이형세포질성변이 한국인의미토콘드리아 DNA에서확인된 52곳의점이형세포질성변이중에서하플로그룹을결정하는주요변이가위치한곳에형성된점이형세포질성변이가 9곳에서확인되었다. 다만이들변이는각변이가확인된개체의하플로그룹이아닌다른계열의하플로그룹을결정하는 64
76 주요변이였다. 각각의변이는다음과같았다. 하플로그룹 B4a1c1a1 의 1 개체에서 93 번째뉴클레오티드 : A&G (A93G = 하플로그룹 B5a2a 를결정하는변이 ) 하플로그룹 B4a1c1a 번째뉴클레오티드 : T&C (T16092C = 하플로그룹 B2o, B2a4 를결정하는변이, 하플로그룹 B4a3, B4c1a2a 를결정하는주요변이중하나 ) 하플로그룹 D4a 의 1 개체에서 번째뉴클레오티드 : T&C (T16126C = 하플로그룹 D5a3a1a 를결정하는주요변이중 하나 ) 하플로그룹 D4a1b 의 1 개체에서 990 번째뉴클레오티드 : T&C (T990C = 하플로그룹 D4h3b 를결정하는주요변이중하나 ) 하플로그룹 G1a1a의 1개체에서 16129번째뉴클레오티드 : A&G (A16129G = 하플로그룹 L 을결정하는주요변이중하나, G16129A! = 하플로그룹 G1b2/G1b3/G1b4 계열을결정하는변이 ) 하플로그룹 G1a1a 의 1 개체에서 번째뉴클레오티드 : A&G (A12361G = 하플로그룹 G1b 를결정하는변이중하나 ) 하플로그룹 G1a1a의 1개체에서 16223번째뉴클레오티드 : T&C (T16223C = 하플로그룹 R을결정하는변이중하나 ), 16234번째뉴클레오티드 : & (C16234T = 하플로그룸 G2a5를결정하는변이중하나 ) 하플로그룹 N9a2a2의 1개체에서 16129번째뉴클레오티드 : G&A (A16129G= 하플로그룹 L 을결정하는변이중하나, G16129A! = 하플로그룹 N9a1 3을결정하는변이, 65
77 G16129A! = 하플로그룹 N9b1b 를결정하는변이중하나 ) 5.4. 세포핵미토콘드리아염기서열 (NUMT) 과점이형세포질성 생거분석이나 VCF 보고서, IGV 등에서섞여있는상태로분석되었더라도 mtdna-server에서해당위치가 NUMT(nuclear mitochondrial sequence; 세포핵미토콘드리아염기서열 ) 의가능성이있는것으로보고한경우에는이형세포질성이아닌것으로간주하였다. 생거분석에서발견된 152번과 146번뉴클레오티드와 IGV에서확인된 204번, 709번, 3150번, 4218번, 9948번, 15261번, 15924번뉴클레오티드가이러한경우에해당되었다. 66
78 고찰 1. 차세대염기서열분석기법에의한미토콘드리아 DNA 전체염기서열분석 1.1 염기서열부위와 coverage 본연구에서차세대염기서열분석기법을적용하여개정케임브리지표준염기서열에따라한국인의미토콘드리아 DNA의전체연기서열을해독할때, 염기서열의부위에따라다른 coverage를보인점은 Ion Torrent PGM 기반의다른연구 (Seo SB, et al., 2015, Zhou Y, et al, 2016) 와유사한양상이었다. Seo 등의연구에서높은 coverage를보였던염기서열부위로서 2,409번에서 3,201번사이의부분과, 낮은 coverage를보였던염기서열부위로서 13,271번에서 14,859번째뉴클레오티드위치를제시했던점, Zhou 등의연구에서낮은 coverage를보였던염기서열부위로서 13600번에서 13800번째뉴클레오티드위치, 14,100번에서 14,900번째뉴클레오티드위치를제시했던점등은, 본연구에서최고높은 coverage를보인염기서열부위로 2,600번에서 2,900번사이, 500배이하의낮은 coverage를보인염기서열부위로 13,600번에서 14,800번사이 (NADH 탈수소효소아단위 5 & 6 유전자부위 ) 였던점과유사하다. 본연구에서뿐만아니라다른연구에서도유사하게나타나는이러한현상은높은 coverage를보였던부위에비해낮은 coverage를보였던부위에서중합체뻗기부위 (polymeric stretch region) 가더많이위치하는등의미토콘드리아 DNA 염기서열부위별특성때문으로생각되었다. (King JL, et al., 2014, Seo SB, et al, 2015, Zhou Y, et al, 2016) ( 표 13 참조 ) 1.2 VCF 의변이검출과확인 VCF 에서보고되지않았거나오검출된변이에있어서, 다른연구와 비교해보았다. ( 표 14 참조 ). 모든연구에서공통적으로 67
79 오검출되었다고했던염기서열부위는 8,281번에서 8,289번째뉴클레오티드위치였다. 중국한족의연구에서 VCF의변이검출보고결과와 IGV로확인한결과와비교하여확인된오검출결과는본연구결과와매우유사하였다. 이들부위들은뉴클레오티드동형중합체 (homopolymer, >2) 를형성하는부위라는점에서, 변이의오검출은이러한특성과관련된것으로생각된다. 본연구에서 9,824번뉴클레오티드위치에서세개의대립형질 (tri-allele) 변이가확인된점에있어서, 중국한족에대한연구에서도동일한결과를제시하였었다. (Parson W, et al, 2013, Seo SB, et al, 2015, Zhou Y, et al, 2016) 조절부위의과변이부위II에서시토신이연이어염기서열을만드는부위 ( 번, 번 ) 에서는시토신삽입을정확하게검출하지못했다. 이는 Parson 등이 Ion Toreent PGM기반의차세대염기서열분석기법에의한분석결과와생거기법에의한염기서열분석결과를비교한연구에서도이부위에대하여시토신삽입에대한불일치가확인되었으며, 다른연구들에서도이부위에대한차세대염기서열분석기법에서는정확하게검출되지않아변이의부위로포함시키지않았다. 뉴클레오티드중합체뻗기부위 (homopolymer stretch region) 및길이이형세포질성에대한분석에있어서 VCF에의한결과에근거한분석은제한점이있는것으로보여서, 이들부위에대해서는 IGV 등으로염기서열을확인하는것이필요하며, 미토콘드리아 DNA 염기서열분석에있어서정렬과알고리듬에있어서이러한미토콘드리아 DNA의특성을반영되어야할것이다. (Parson W, et al., 2013, Seo SB, et al., 2015, Zhou Y, et al., 2016) 2. 한국인에서미토콘드리아 DNA 의유전적특징 2.1 한국인에서변이의분포와양상 본연구에서 186명의한국인을대상으로한전체염기서열분석결과 455곳의뉴클레오티드위치에서 6,495개의변이가관찰되었으며, 평균 1개체당 34.9개의변이가있는것이었다. 조절부위에서 1,682개 (25.9%), 암호부위에서 4,813개 (74.1%) 가관찰되었으며, 이중대부분의변이는치환으로서 6,336개 (97.55%), 나머지는삽입결실이 68
80 159 개 (2.45%) 관찰되었던점을다른민족과함께비교해보았다. ( 표 15 참조 ) 중국한족 107명의한족을대상으로하였으며 725곳의뉴클레오티드위치에서 4,022개의변이가관찰되었고, 이는평균 1개체당 37.6개의변이가관찰되는것이었다. 조절부위에서관찰된변이는 892개 (22.18%) 였다. 변이의구성으로는치환에의한변이가 3,919개 (97.44%), 삽입결실에의한변이가 103개 (2.56%) 였다. (Zhou Y, et al., 2016) 에스토니아인 114명의에스토니아인을대상으로하였으며, 512곳의뉴클레오티드위치에서 2,663의변이가관찰되었으며, 이는평균 1개체당 23.4개의변이가관찰되는것이었다. 607개 (22.8%) 는조절부위에서, 나머지 2,056개 (77.2%) 는암호부위에서관찰되었다. (Stoljarova M, et al., 2016) 미국인 ( 아프리칸미국인, 코카시안인, 히스패닉인 ) 283명의미국인으로 87명의아프리칸미국인, 83명의코카시안인, 113명의히스패닉인을대상으로하였다. 11,607개의변이가 1,353곳의뉴클레오티드위치에서관찰되었고, 이는 1개체당평균 41.0개의변이가관찰되었다. 8,669개 (74.7%) 의변이가암호부위에서관찰되었다. (King JL, et al., 2014) 조절부위와암호부위에서관찰되는변이의빈도는여러민족에서유사하게조절부위의변이가 22 25% 범위내에서관찰되는양상을보임을알수있었다. 한국인과비교하였을때, 여러인종을포함하는미국인의경우더많은곳의뉴클레오티드위치에서변이가관찰되었으며, 1개체당관찰되는평균변이의수도가장많았다. 동아시아안에서거주하는중국의한족과한국인을같이비교하였을때, 변이의구성이나분포가서로유사한양상을보이고있음을알수있었다. ( 표 15 참조 ) 2.2 한국인에서공통변이 69
81 한국인에서나타나는고유의유전적다양성을알아보기위해 186명의한국인전체에서공통으로나타나는변이와절반이상에서관찰되는변이들을개정캠브리지표준서열에근거하여분석하고이를같은동아시아에있는중국의한족과일본인과비교해보았다. 일본인의경우조절부위에대한분석자료만가능해서비교분석에제한은있었으나, 한국인, 중국한족, 일본인에서유사한경향을보임을알수있었다. 다만동아시아이외에에스토니안인과미국인을포함하여분석하였을때, 모든개체에서공통적으로나타나는변이의경우상당한부분의변이는다른민족에서도나타나기때문에이들변이들이각민족의유전적특성으로보기는어려울것으로생각되었다. 각민족에서절반이상의개체에서나타난변이의경우에는한국인과중국한족은거의동일한양상을보이지만그외다른민족과의비교에서는다른양상을보이고있음을알수있다. (Behar DM, et al., 2012, King JL, et al., 2014, Maruyama S, et al., 2013, Stoljarova M, et al., 2016, van Oven M, et al., 2009, Zhou Y, et al., 2016) ( 표 16 참조 ) 2.3 한국에서하플로그룹의세분화양상 미토콘드리아 DNA 조절부위로만근거하였을때 186명의한국인에서 14개의하플로그룹 ( 조절부위 ) 은, 전체염기서열분석결과 47개의하플로그룹으로세분화될수있었다. 조절부위에근거하여한국인에서흔히나타나는하플로그룹 D, A5a, M7, G1a1, B4, N9a2a, 세분화가안되어큰군으로묶였던하플로그룹 M9;G;D, 제주도에서높은빈도로나타나는하플로그룹 Y1에대하여전체염기서열분석후세분화되는양상을분석해보았다. 하플로그룹 D4의 4개의하플로그룹이 8개의하위하플로그룹으로세분화되었고, 하플로그룹 M9;G;D는모두 6개의하플로그룹 D의하위하플로그룹으로재분류되었다. 이들군의경우암호부위와조절부위의변이의비를보면각각 3.48, 4.59로, 전체변이중암호부위에있는변이가훨씬많은부분을차지하고있다. 따라서암호부위변이분석을포함하는전체염기서열분석결과가하플로그룹을결정하는데더정확한정보를줄수있음을알수있었다. 하플로그룹 A5a군도 5개의하위하플로그룹으로, 2개의하플로그룹 M7은모두 7개의하위하플로그룹으로, 하플로그룹 F군은 6개의하위하플로그룹으로분류될수있었다. 70
82 하플로그룹 B군의경우, 암호부위와조절부위의변이의비가 1.51로 다른하플로그룹에비해낮게나타나고, 조절부위를근거로하였을때 하플로그룹이 하위 하플로그룹으로 세분화되지 못하고 상위 하플로그룹으로 묶여서 보여줄 뿐만 아니라 하플로그룹 분석툴 (MitoTool, Haplogrep) 에따라하플로그룹분류에차이가나는결과를 보여주었으나, 전체염기서열분석결과는 명확하게 하위 하플로그룹으로 세분화시켜주었다. 대조적으로 하플로그룹 G1a1의 경우 약절반정도에서 1개의 하위 하플로그룹을추가하여세분화할수있었고, 하플로그룹 N9a2a의경우 1개의 하위 하플로그룹이 추가적으로 세분화 될 수는 있었으나 대부분은 그대로 하플로그룹 N9a2a로 변화가 없었다. 하플로그룹 Y1군의 경우도 3군의 하위 하플로그룹을 추가적으로 세분화할 수 있었지만대부분은그대로 Y1군으로변화가없었다. 하플로그룹 Y1의경우제주도에서흔히관찰되는유형이다. 제주도는 작고고립된인구를가진곳으로오랜세월동안다른지역인구와의 교류가 제한되었던 곳이다. 따라서 이 인구에 속한 각 개체들은 유전적으로 서로 밀접하게 연관되어 있을 것으로 기대되었기에, 미토콘드리아 DNA 전체염기서열분석결과확인된암호부위변이로도 하플로그룹 Y1의각개체를식별하는것은여전히쉽지않았다. 본연구에서 분석한 한국인의 하플로그룹의 양상은 일본인과 중국 한족과 유사하였다. 일본인에서 하플로그룹 D4가 가장 높은 빈도로 관찰되었다. 107명의 한족에서 60개의 하플로그룹이었으나 전체염기서열로 근거하였을 때 74개의 하플로그룹으로 세분화되었다. 한족에서나타난주요하플로그룹으로는하플로그룹 M, D, F, B, C, A, N 등으로, 매크로하플로그룹 M( 하플로그룹 D, G, M) 이 약 48% 를 차지하고 있던 점은 한국인에서도 매크로하플로그룹 M이 약 57% 였음과 유사한 결과였다. 또한 조절부위에 근거하였을 때보다 전체염기서열에 근거하였을 때, 하플로그룹이 더 세분화되는 경향도 유사하게나타났다. 본연구에서조절부위만으로하였을때는하플로그룹 M9;G;D로나타나서세분화되지못했던군이전체염기서열분석후에는 하플로그룹 D의 하위 하플로그룹으로 재분류되었던 것과 유사하게 한족에서도조절부위에근거했을때는하플로그룹 M으로나타났던것이 전체염기서열분석후에는하플로그룹 D와 G 등의하위하플로그룹으로 다시 결정되는 유사한 양상이 관찰되었다. [ 표 17, 표 18 참조 ] (Maruyama S, et al., 2013, Zhou Y, et al., 2016) 71
83 미국인 283명의개체에대한전체염기서열분석결과전체 208개의하플로그룹이확인되었고, 각인종별로세분화하여보면아프리칸 (87명), 코카시안 (83명), 히스패닉 (113명) 에서각각 70개, 79개, 70개의하플로그룹으로세분화되었다. 아프리칸미국인에서주요하플로그룹으로하플로그룹 L이었고, 코카시안미국인의경우하플로그룹 H, U, T, K 등이있었으며, 히스패닉미국인의경우하플로그룹 A, C, B, H, L, U, D, T 등이있었다. 에스토니안의경우 114명에서 87개의하플로그룹으로세분화되었다. 주요하플로그룹으로는 D, HV, I, J, M, N, R, T, U, W, X 등으로하플로그룹 HV가약 47%, 하플로그룹 U가약 24% 를차지하고있었고, 하플로그룹 D의경우 1례가있었다. 이러한결과는한국인과중국한족에서주로나타나는하플로그룹과다른양상으로나타남을알수있었다. (King JL, et al., 2014, Maruyama S, et al., 2013, Stoljarova M, et al., 2016, Zhou Y, et al., 2016) 조절부위를근거했을때와전체염기서열을근거하였을때새로운하플로그룹으로재분류되는현상은본연구와중국한족뿐만아니라다른연구에서도유사하게관찰되었다. ( 표 18 참조 ) 본연구에서는하플로그룹 M9;G;D( 전체의약 9%) 에서하플로그룹의재분류가있었고, 한족의연구에서는약 6.5%, 미국인에서는 3.2%, 에스토니안연구에서 0.8%(1개체 ) 에서있었다. 재분류되는양상은다른계통의하플로그룹으로재분류되거나, 같은계통이나세분화가아닌다른하위하플로그룹으로재분류되는현상으로나타났다. 특히하플로그룹 D, G, M에서는공통적으로이러한재분류가있었다. 따라서하플로그룹을결정할때, 특히동아시아에서는매크로하플로그룹 M이주요하플로그룹이라는점을감안한다면, 조절부위에근거한하플로그룹분석자료는유전적다양성에대한분석에제한점이있을수있고, 전체염기서열을고려해야할것으로생각된다. (King JL, et al., 2014, Maruyama S, et al., 2013, Stoljarova M, et al., 2016, Zhou Y, et al., 2016) 2.4 한국인에서나타나는하플로타입 (haplotype) 의양상 전체염기서열분석결과 하플로타입의 개수가 증가하여 개체수에 근접하는경향을보여주었고, 이러한경향은다른연구와유사하였다. ( 표 10, 그림 7 참조 ) 본 연구에서 186명의 개체에서 조절부위에 72
84 근거하였을때확인된 70개의하플로타입이 137개의하플로타입으로관찰되었다. 이는중국한족의연구에서도유사한경향을보여주었는데, 107명의개체에서조절부위로근거하였을때 103개의하플로타입이었고전체염기서열분석에서는 107개로나타났다. 미국인 (588명, 아프리칸 170명, 코카시안 263명, 히스패닉 155명 ) 에대하여생거기법으로전체염기서열을분석한연구에서, 조절부위에근거하였을때하플로타입이 470개 ( 아프리칸 130, 코카시안 211, 히스패닉 130) 였던것이, 전체염기서열에근거하였을때하플로타입은 575개 ( 아프리칸 169, 코카시안 259, 히스패닉 147) 였다. 또한다른연구에서도전체염기서열분석결과개체수에근접한하플로그룹개수를보여주었는데, 미국인에대한다른연구에서 283명의개체에대하여전체염기서열분석결과 279개의하플로타입이확인되었고, 세부적으로보면아프리칸미국인 (87명), 코카시안미국인 (83명), 히스패닉미국인 (113명) 에서하플로타입은각각 85, 83, 111개으로나타났다. 에스토니안에대한연구에서도 114명의개체에서 100개의하플로타입이확인되었다. (Just RS, et al., 2015, King JL, et al., 2014, Maruyama S, et al., 2013, Stoljarova M, et al., 2016, Zhou Y, et al., 2016) 2.5 한국인에서나타나는개별변이양상 본연구에서하플로그룹결정에기여하지않으면서도같은하플로그룹임에도서로다른변이들이관찰되었다. 이러한변이들은다른민족의연구에서도관찰되는현상이었다. 하나의개체에서만발견되는변이라면개체의자발적돌연변이로여겨질수있겠으나, 2개체이상에서발견되는변이라면그러한변이에의해새로운계열로정의될수있는근거가될수있을것이다. 특히한국인의연구에서다른민족과비교해보았을때, 하플로타입은개체수에근접한개수를보여주면서도, 결정된하플로그룹의수는개체수에비해적은점이관찰되었다. 이는한국인에서잠재적으로새로운하위하플로그룹의가능성을시사하는소견일수도있다. 그러나이들개체사이에서로친족관계가없음에도새로운변이를공통으로가지고있다고관찰되었을지라도, 새로운세부군으로간주하기위해서는그들사이공통적인조상에대한정보에대한분석이필요할것이며, 또한변이의관찰이반복적으로관찰된다고하는것이확인되는등더많은연구가 73
85 필요할것이다. (Kong QP, et al., 2010, Maruyama S, et al., 2013, van Oven M, 2010) ( 표 7 참조 ) 3. 한국인에서미토콘드리아 DNA 점이형세포질성 점이형세포질성을결정하는기준에있어서부요소를인지하는최소수준 (threshold) 으로서대개 10% 가회자되고는있으나연구마다이형세포질성에대하여해석하는기준이조금씩상이하여, 부요소의최소기준의정의에서부터최소대립형질의뉴클레오티드의빈도, 최소 coverage, 주변의연속하는 3염기에삽입결실이없는경우, 한개체에여러개의이형세포질성변이가관찰되면서새로운하플로그룹을형성하는등의여러엄격한기준을적용한정의에이르기까지다양한수준과범위의적용기준이있었다. ( 표 19 참조 ) 본연구에서는부요소에대하여최소 40배이상의 coverage를보여주면서세포핵에전위된미토콘드리아염기서열 (numt) 도제외시키고, mtdna-server을통하여신뢰할수있는자료인지입증된결과를이형세포질성변이로관찰하였다. 다른연구에서발표된자료와본연구의결과를직접적으로비교하는것은제한점은있으나, 본연구결과의점이형세포질성변이에대한결과를다른연구와비교하였을때전반적으로유사한양상을보여주었다. [ 표 19] (Holland MM, et al., 2011, Just RS, et al., 2015, Li M, et al., 2010, Skonieczna K, et al., 2015, Sosa MX, et al., 2012, Weissensteiner H, et al., 2016, Zhou Y, et al., 2016) 본연구에서는각개체의혈액에서미토콘드리아 DNA 염기서열에서나타나는이형세포질성변이에대해서분석하였다. 동일개체의여러장기에서미토콘드리아 DNA 염기서열에대하여분석한다른연구와비교하였을때본연구에서처럼이형세포질성변이가염기서열에서고르지않고특정부위들에몰려나타나는현상은유사하게관찰되었다. 다만이전연구에따르면, 특정장기, 특정대립형질뉴클레오티드, 연령에따라점이형세포질성의분포가다르게나타나고, 점이형세포질성의빈도는연령, 특정대립형질뉴클레오티드와관련이있다고하였다. 한국인에서점이형세포질성변이의양상과특성을파악하는데있어서, 동일개체내여러장기및혈액에서나타나는이형세포질성의분포와차이, 이형세포질성변이의호발부위및특정 74
86 대립형질뉴클레오티드, 연령분석등에대한연구가필요할것이다. (Li M, et al., 2015) 4. 한국인에서미토콘드리아 DNA 전체염기서열 분석결과와개체식별력 전체염기서열분석을통해개체식별력을높일수있음을확인할수 있었다. 한국인에서매크로하플로그룹 M(M9;G;D, D, G, M 등 ) 은주요 하플로그룹이면서, 조절부위 변이로는 하위 하플로그룹 유형 분류에 제한적인 결과를 보여주었기에, 전체염기서열분석을 통해 이들 하플로그룹의하위하플로그룹유형결정이가능할것으로기대하였었다. 연구결과, 공통의하플로그룹에서하위하플로그룹의분류를위해서는 전체염기서열분석을통해조절부위뿐만아니라암호부위에서나타나는 변이에대한정보는하플로그룹세분화및재분류에기여하였을뿐만 아니라, 하플로그룹에 기여하지 않지만 개체간 차이를 만들어내는 개별변이를확인할수있었다. 하플로타입에있어서더증가된개수의 고유의 하플로타입을 확인할 수 있었으며 이에 대한 식별력 (discriminating capacity) 역시 전체염기서열분석결과 증가한 것을 확인 할 수 있었다 ( 표 10, 그림 8, 그림 9 참조 ). 따라서 전체염기서열분석은 개체식별에 더 많은 정보를 줄 수 있음을 확인하였다. 또한 조절부위에 근거한 하플로그룹이 전체염기서열분석결과새로운하플로그룹으로재분류될수있는점을 감안하였을때, 조절부위에근거하여분류된하플로그룹의정보의경우 개체식별해석에있어서주의가필요할것으로생각된다. (King JL, et al., 2014, Maruyama S, et al., 2013, Stoljarova M, et al, 2016, Tanaka M, et al, 2004, Zhou Y, et al., 2016) 중국의 한족이나 미국인, 에스토니안인에 대한 연구에서 전체염기서열분석결과 결정된 하플로그룹의 개수와 개체수의 비율은 69 80% 를 차지하지만, 한국인에서는 25% 로 비교적 낮았다. 하플로타입에서는 한국인 (73.7%) 과 다른 민족 (88 99%) 과 거의 유사하였다. 또한하플로그룹 N과 Y의경우전체염기서열분석결과에도 대부분은하플로그룹분류에변화가없지만하플로타입에서는개체수에 근접한 서로 다른 유형이 관찰되었다. 이러한 양상이 한국인에서의 75
87 유전적특성일수도있으나아마도본연구에서해당실험대상군의작은규모때문일수도있을것이다. 따라서앞으로의연구에서더많은한국인을대상으로분석할필요가있을것이다. (King JL, et al., 2014, Maruyama S, et al., 2013, Stoljarova M, et al, 2016, Tanaka M, et al, 2004, Zhou Y, et al., 2016) 개체식별에서있어서한국인에서나타나는이형세포질성의특성등에대한정보가필요하며, 해석에주의하여야할것이다. 본연구에서확인된 52곳의점이형세포질성중 3곳의점이형세포질성 (B4a1c1a np8850, D4a1 np16103, D4a1c np4088) 은고유의하플로타입을결정함에있어서유일한차이를보인변이여서별개의하플로타입으로구분하기어려웠다. 또한동일개체서로다른여러장기에서미토콘드리아 DNA염기서열을분석한연구에서여러장기에서공통적으로나타나는이형세포질성변이는유전성변이로볼수있겠으나특정장기에서만나타난다면체세포돌연변이일것이므로, 이러한정의로접근하였을때, 관찰된이형세포질성변이의 43-65% 는유전성변이라고하였다. 본연구에서 186명의한국인중 40개체 (21.51%) 에서이형세포질성이관찰되었는데, 혈액등의 1종류의시료에서실험이진행되었고, 실험군에대한가계도나유전적정보는알수없었기때문에그이상의분석은어려웠다. 가계도나유전관계를알수있고다양한연령대를포함하는한국인을대상으로하여유전성변이와체세포돌연변이의분포와양상, 연령에의한이형세포질성변이의변화및양상에대한연구가필요할것이다. (Li M, et al., 2015) 76
88 결론 본연구에서한국인에서흔하게나타나는하플로그룹을중심으로 186명의한국인을선별하여차세대염기서열분석기법을적용하여미토콘드리아 DNA 전체염기서열분석을시행하여한국인에서나타나는미토콘드리아 DNA의유전적특성을파악할수있었다. 미토콘드리아 DNA 전체염기서열의조절부위와암호부위에서나타나는변이의분포는다른인종과유사하였지만, 변이의위치와구성은다른인종과차이가있었지만동아시아인특히중국한족과는유사하였다. 하플로그룹및하플로타입에있어서전체염기서열분석결과조절부위만을근거로하였을때보다하플로그룹의세분화및재분류, 증가된개수의하플로타입이확인되었으며, 이러한경향은다른인종에서의전체염기서열분석결과와유사하였다. 각하플로그룹의양상은중국의한족과유사하였다. 점이형세포질성의경우, 각이형세포질성의분포와특성은다른인종을대상으로한이형세포질성연구와유사한양상을보였다. 한국인에서개체식별력에있어서는조절부위만을근거로하였을때보다하플로그룹이세분화및재분류되고, 개체수에근접한하플로타입이결정되어개체식별력을높여줄수있음이확인되었다. 이러한한국인의미토콘드리아 DNA의유전적다양성은한국인의개체식별에있어서모계분석및여러민족간구분에서중요한기초자료가될뿐만아니라한국인에서여러주요질병과미토콘드리아 DNA의변이또는하플로그룹과의연관성에대한연구에있어서기초자료가될것으로본다. 본연구결과가앞으로한국인에서미토콘드리아 DNA의유전적다양성및질병연구에중요한견인차역할을할것으로기대한다. 77
89 참고문헌 Akhmedov AT. Marín-García J. Mitochondrial DNA maintenance: an appraisal. Mol Cell biochem 2015;409: Alberts B, Johnson A, Lewis J. Raff M, Roberts K, Walter P. Molecular biology of the cell, 5th ed. New York: Garland Science; 2008, p Alberts B, Johnson A, Lewis J. Raff M, Roberts K, Walter P. Molecular biology of the cell, 5th ed. New York: Garland Science; 2008, p Allard MW, Wilson MR, Monson KL, Budowle B. Control region sequences for East Asian individuals in the Scientific Working Group on DNA Analysis Methods forensic mtdna data set. Leg Med (Tokyo) 2004;6: Anderson S, Bankier AT, Barrell BG, de Bruijin MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, Schreier PH, Smith AJ, Staden R, Young IG. Sequence and organization of the human mitochondrial genome. Nature 1981;290: Andrews RM, Kubacka I, Chinnery PF, Lightowlers RN, Turnbull DM, Howell N. Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA. Nat Genet 1999;23:147. Ayala FJ. The myth of Eve: molecular biology of human origins. Science 1995;270: Bandelt HJ, Lahermo P, Richards M, Macaulay V. Detecting errors in mtdna data by phylogenetic analysis. Int J Legal Med 2001;115:
90 Bandelt HJ, Macaulay V, Richards M. Human mitochondrial DNA and the evolution of Homo sapiens. 1 st ed. Germany: SpringerBerlin Heidelberg New York; 2006, Behar DM, van Oven M, Rosset S, Metspalu M, Loogväli EL, Silva NM, Kivisild T, Torroni A, Villems R. A Copernican Reassessment of the human mitochondrial DNA tree from its root. Am J Hum Genet 2012;90: Bendall KE, Macaulay VA, Baker JR, Sykes BC. Heteroplasmic point mutations in the human mtdna control region. Am J Hum Genet 1996;59: Bogenhagen D, Clayton DA. Mouse L cell mitochondrial DNA molecules are selected randomly for replication throughout the cell cycle. Cell 1977;11: Bogenhagen DF. Mitochondrial DNA nucleoid structure. Biochim Biophys Acta 2012;1819: Brown WM, Prager EM, WangA, Wilson AC. Mitochondrial DNA sequence of primates: tempo and mode of evolution. J Mol Evol 1982;18: Budowle B, Allard MW, Wilson MR, Chakraborty R. Forensics and mitochondrial DNA: applications, debates, and foundations. Annu Rev Genomics Hum Genet 2003;4: Cann RL, Stoneking M, Wilson AC. Mitochondrial DNA and human evolution. Nature 1987;325:31-6. Chomyn A, Cleeter MW, Ragan CI, Riley M, Doolittle RF, Attardi G. URF6, last unidentified reading frame of human mtdna, codes for an NADH dehydrogenase subunit. Science 1986;234:
91 Choung CM, Jang BS, Li Z, Zhang A, Lee YS, Lee JB, Lee SD. Heteroplasmy pattern within D-loop region of mitochondrial DNA in Koreans. Kor J Legal Med 2005;29: Clayton DA. Structure and function of the mitochondrial genome. J Inher Metab Dis 1992;15: Coble MD, Just RS, O'Callaghan JE, Letmanyi IH, Peterson CT, Irwin JA, Parsons TJ. Single nucleotide polymorphisms over the entire mtdna genome that increase the power of forensic testing in Caucasians. Int J Legal Med 2004;118: Coble MD, Loreille OM, Wadhams MJ, Edson SM, Maynard K, Meyer CE, Niederstätter H, Berger C, Berger B, Falsetti AB, Gill P, Parson W, Finelli LN. Mystery solved: the identification of the two missing Romanov children using DNA analysis. PLoS One 2009;4:e4838. Elson JL, Lightowlers RN Mitochondrial DNA clonality in the dock: can surveillance swing the case? Trends Genet 2006;22: Fan L, Yao YG. An update to MitoTool: using a new scoring system for faster mtdna haplogroup determination. Mitochondrion 2013;13: Fendt L, Zimmermann B, Daniaux M, Parson W. Sequencing strategy for the whole mitochondrial genome resulting in high quality sequences. BMC Genomics 2009;10:139. Garrigan D, Hammer MF. Reconstructing human origins in the genomic era. Nature Rev Genet 2006;7: Gibbons A. Mitochondrial Eve: Wounded, but not dead yet. Science 1992;257:
92 Giles RE, Blanc H, Cann HM, Wallace DC. Maternal inheritance of human mitochondrial DNA. Proc Natl Acad Sci USA 1980;77: Gonder MK, Mortensen HM, Reed FA, de Sousa A, Tishkoff SA. Whole-mtDNA genome sequence analysis of ancient African lineages. Mol Biol Evol 2007;24: Greenberg. BD, Newbold JE, Sugino A. Intraspecific nucleotide sequence variability surrounding the origin of repolication in human mitochondrial DNA. Gene 1983;21: Hasegawa M, Di Rienzo A, Kocher TD, Wilson AC. Toward a more accurate time scale for the human mitochondrial DNA tree. J Mol Evol 1993;37: Hauswirth WW. Laipis PJ. Mitochondrial DNA polymorphism in a maternal lineage of Holstein cows. Proc Natl Acad Sci USA 1982;79: Hayashida K, Omagari K, Masuda JI, Hazama H, Kadokawa Y, Ohba K, Kohno S. The sperm mitochondria-specific translocator has a key role in maternal mitochondrial inheritonce. Cell Biol int 2005;29: Heather JM, Chain B. The sequence of sequencers: The history of sequencing DNA. Genomics 2016;107:1-8. Holland MM, Parsons TJ. Mitochondrial DNA Sequence Analysis - Validation and Use for Forensic Casework. Forensic Sci Rev 1999;11: Holland MM, McQuillan MR, O'Hanlon KA. Second generation sequencing allows for mtdna mixture deconvolution and high resolution detection of heteroplasmy. Croat Med J 2011;52:299-81
93 313. Hong SB, Kim KC, Kim W. Mitochondrial DNA haplogroups and homogeneity in the Korean population. Genes Genom 2014;36: Jin HJ, Kwak KD, Hong SB, Shin DJ, Han MS, Tyler-Smith C, Kim W. Forensic genetic analysis of mitochondrial DNA hypervariable region I/II sequences: an expanded Korean population database. Forensic Sci Int 2006;158: Jin HJ, Tyler-Smith C, Kim W. The Peopling of Korea Revealed by Analysed of Mitochondrial DNA and Y-Chromosomal Markers. PLos One 2009;4:e4210. Just RS, Scheible MK, Fast SA, Sturk-Andreaggi K, Röck AW, Bush JM, Higginbotham JL, Peck MA, Ring JD, Huber GE, Xavier C, Strobl C, Lyons EA, Diegoli TM, Bodner M, Fendt L, Kralj P, Nagl S, Niederwieser D, Zimmermann B, Parson W, Irwin JA. Full mtgenome reference data: development and characterization of 588 forensic-quality haplotypes representing three U.S. populations. Forensic Sci Int Genet 2015a;14: Just RS, Irwin JA, Parson W. Mitochondrial DNA heteroplasmy in the emerging field of massively parallel sequencing. Forensic Sci Int Genet 2015b;18: King JL, LaRue BL, Novroski NM, Stoljarova M, Seo SB, Zeng X, Warshauer DH, Davis CP, Parson W, Sajantila A, Budowle B. Highquality and high-throughput massively parallel sequencing of the human mitochondrial genome using the Illumina MiSeq. Forensic Sci Int Genet 2014;12: Kivisild T, Tolk HV, Parik J, Wang Y, Papiha SS, Bandelt HJ, Villems R. The emerging limbs and twigs of the East Asian mtdna 82
94 tree. Mol Biol Evol 2002;19: Koehler CM, Lindberg GL, Brown DR, Beitz DC, Freeman AE, Mayfield JE, Myers AM. Replacement of bovine mitochondrial DNA by a sequence variant within one generation. Genetics 1991;129: Kong QP, Yao YG, Sun C, Bandelt HJ, Zhu CL, Zhang YP. Phylogeny of east Asian mitochondrial DNA lineages inferred from complete sequences. Am J Hum Genet 2003;73: Kong QP, Yao YG, Liu M, Shen SP, Chen C, Zhu CL, Palanichamy MG, Zhang YP. Mitochondrial DNA sequence polymorphisms of five ethnic populations from northern China. Hum Genet 2003;113: Kong QP, Bandelt HJ, Zhao M, Zhang Y, Yao Y. Reply to van Oven: suggestion and caveats for naming mtdna haplogroup. Proc Natl Acad Sci USA. 2010;107:E40-1. Lee HY, Yoo JE, Park MJ, Chung U, Kim CY, Shin KJ. East Asian mtdna haplogroup determination in Koreans: haplogroup-level coding region SNP analysis and subhaplogroup-level control region sequence analysis. Electrophoresis 2006;27: Lee HY, Yoo JE, Park MJ, Chung U, Shin KJ. Mitochondrial DNA control region sequences in Koreans: identification of useful variable sites and phylogenetic analysis for mtdna data quality control. Int J Legal Med 2006;120:5-14. Lee SD, Lee YS, Lee JB. Polymorphism in the mitochondrial cytochrome B gene in Koreans. An additional marker for individual identification. Int J Legal Med 2002;116:74-8. Li M, Schönberg A, Schaefer M, Schroeder R, Nasidze I, Stoneking 83
95 M. Detecting heteroplasmy from high-throughput sequencing of complete human mitochondrial DNA genomes. Am J Hum Genet 1020;87: Li M, Schrönder R, Shengyu Ni, Madea B, Stoneking M. Extensive tissure-related and allele-related mtdna heteroplasmy suggests positive selection for somatic mutations. Proc Natl Acad Sci USA 2015;112: Lopez JV, Yuhki N, Masuda R, Modi W, O Brien SJ. Numt, a recent transfer and tandem amplification of mitochondrial DNA to the nuclear genome of the domestic cat. J Mol Evol 1994;39: Lutz-Bonengel S, Schmidt U, Schmitt T, Pollak S. Sequence polymorphisms within the human mitochondrial genes MTATP6, MTATP8, and MTND4. Int J Legal Med 2003;117: Maruyama S, Komuro T, Izawa H, Tsutsumi H. Analysis of human mitochondrial DNA polymorphisms in the Japanese population. Biochem Genet 2013;51: Metzker ML. Sequencing technologies the next generation. Nat Rev Genet 2010;11: Moraes CT, Srivastava S, Kirkinezos I, Oca-Cossio J, van Waveren C, Woischnick M, Diaz F. Mitochondrial DNA structure and function. Int Rev Neurobiol 2002;53:3-23. Nilsson M, Andréasson-Jansson H, Ingman M, Allen M. Evaluation of mitochondrial DNA coding region assays for increased discrimination in forensic analysis. Forensic Sci Int Genet 2008;2:1-8. Parsons TJ, Muniec DS, Sullivan K, Woodyatt N, Alliston-Greiner R, Wilson MR, Berry DL, Holland KA, Weedn VW, Gill P, Holland MM. 84
96 A high observed substitution rate in the human mitochondrial DNA control region. Nat Genet 1997;15: Parsons TJ, Holland MM. Mitochondrial mutation rate revisited: hot spots and polymorphisms Reply. Nat Genet 1998;18: Parsons TJ, Coble MD. Increasing the forensic discrimination of mitochondrial DNA testing through analysis of the entire mitochondrial DNA genome. Croat Med J 2001;42: Parson W, Strobl C, Huber G, Zimmermann B, Gomes SM, Souto L, Fendt L, Delport R, Langit R, Wootton S, Lagacé R, Irwin J. Evaluation of next generation mtgenome sequencing using the Ion Torrent Personal Genome Machine (PGM). Forensic Sci Int Genet 2013;7: Parson W, Gusmão L, Hares DR, Irwin JA, Mayr WR, Morling N, Pokorak E, Prinz M, Salas A, Shneider PM, Parsons TJ. DNA commission of the international society for forensic genetics; revised and extended guidelines for mitochondrial DNA typing. Forensic Sci Int Genet. 2014;13: Parson W, Huber H, Moreno L, Madel MB, Bradhagen MD, Nagl S, Xavier C, Eduardoff M, Callaghan TC, Irwin JA. Massively parallel sequencing of complete mitochondrial genomes from hair shaft samples. Forensic Sci Int Genet 2015;15:8-15. Poulton J, Turnbull DM. 74 소 Enmc international workshop; mitochondrial diseases. Neuromuscul Disord 2000;10: Quail MA, Smith M, Coupland P, Otto TD, Harris SR, Connor TR, Bertoni A, Swerdlow HP, Gu Y. A tale of three next generation sequencing platforms: comparison of Ion Torrent, Pacific Biosciences and Illumina MiSeq sequencers. BMC Genomics 2012;13:
97 Redd AG, Takezaki N, Sherry ST, McGarvey ST, Sofro ASM, Stoneking M. Evolutionary history of the COII/tRNALys intergenic 9 base pair deletion in human mitochondrial DNAs from the Pacific. Mol Biol Evol 1995;12: Richards MB, Macaulay VA, Bandelt HJ, Sykes BC. Phylogeography of mitochondrial DNA in western Europe. Ann Hum Genet 1998;62: Rosenthal CK, Heinrichs A, Gray N, Rusk N, Baumann Kim, van Ooij C, Rosenthal CK, Schuldt A, Chenette EJ, Doerr A, Grisendi S, Tonzani S, Whalley K, Le Bot N, David R, Evanko D, Swaminathan S, Wrighton KH, Cesari F, de Souza N. Nature milestones in Light microscopy Macmillan Publishers S6-S22. Rothberg JM, Hinz W, Rearick TM, Schultz J, Mileski W, Davey M, Leamon JH, Johnson K, Milgrew MJ, Edwards M, Hoon J, Simons JF, Marran D, Myers JW, Davidson JF, Branting A, Nobile JR, Puc BP, Light D, Clark TA, Huber M, Branciforte JT, Stoner IB, Cawley SE, Lyons M, Fu Y, Homer N, Sedova M, Miao X, Reed B, Sabina J, Feierstein E, Schorn M, Alanjary M, Dimalanta E, Dressman D, Kasinskas R, Sokolsky T, Fidanza JA, Namsaraev E, McKeman KJ, Williams A, Roth GT, Bustillo J. An integrated semiconductor device enabling non-optical genome sequencing. Nature 2011;475: David C. Samuels,1 Andrew D. Carothers,2 Robin Horton,3 and Patrick F. Chinnery. The power to detect Disease associations with mitochondrial DNA haplogroups. Am J Hum Genet 2006;78: Sato M. Sato K. Maternal inheritance of mitochondrial DNA by diverse mechanisms to eliminate paternal mitochondrial DNA. Biochim Biophys Acta 2013;1833: Scheffler IE. Mitochondria. 2nd ed. Hoboken, New Jearsy. John 86
98 Wiley & Sons, Inc, Hoboken, New Jersey Schwart M, Vissing J. Paternal inheritance of mitochondrial DNA. N Engl J Med 2002;347: Seo SB, Zeng X, King JL, Larue BL, Assidi M, Al-Qahtani MH, Sajantila A, Budowle B. Underlying Data for Sequencing the Mitochondrial Genome with the Massively Parallel Sequencing Platform Ion Torrent PGM. BMC Genomics 2015;16 Suppl 1:S4. Seo Y, Uchiyama T, Shimizu K, Takahama K. Identification of remains by sequencing of mitochondrial DNA control region. Am J Forensic Med Pathol 2000;21: Shendure J, Mitra RD, Varma C, Church GM. Advanced sequencing technologies: methods and goals. Nat Rev Genet 2004;5: Schendure J, Ji H. Next-generation DNA sequencing. Nat Biotechnol 2008;26: Skonieczna K, Malyarchuk B, Jawień A, Marszałek A, Banaszkiewicz Z, Jarmocik P, Borcz M, Bała P, Grzybowski T. Heteroplasmic substitutions in the entire mitochondrial genomes of human colon cells detected by ultra-deep 454 sequencing. Forensic Sci Int Genet 2015;15: Sosa MX1, Sivakumar IK, Maragh S, Veeramachaneni V, Hariharan R, Parulekar M, Fredrikson KM, Harkins TT, Lin J, Feldman AB, Tata P, Ehret GB, Chakravarti A. Next-generation sequencing of human mitochondrial reference genomes uncovers high heteroplasmy frequency. PLoS Comput Biol 2012;8:e Stewart JB, Chinnery PF. The dynamics of mitochondrial DNA heteroplasmy: implications for human health and disease. Nat Rev Genet 2015;16:
99 Stoljarova M, King JL, Takahashi M, Aaspõllu A, Budowle B. Whole mitochondrial genome genetic diversity in an Estonian population sample. Int J Legal Med 2016;130: Tanaka M, Cabrera VM, González AM, Larruga JM, Takeyasu T, Fuku N, Guo LJ, Hirose R, Fujita Y, Kurata M, Shinoda K, Umetsu K, Yamada Y, Oshida Y, Sato Y, Hattori N, Mizuno Y, Arai Y, Hirose N, Ohta S, Ogawa O, Tanaka Y, Kawamori R, Shamoto-Nagai M, Maruyama W, Shimokata H, Suzuki R, Shimodaira H. Mitochondrial genome variation in Eastern Asia and the peopling of Japan. Genome Res 2004;14: Templeton JE, Brotherton PM, Llamas B, Soubrier J, Haak W, Cooper A, Austin JJ. DNA capture and next-generation sequencing can recover whole mitochondrial genomes from highly degraded samples for human identification. Investig Genet 2013;4:26. Titford M. The long history of hematoxylin. Biotech Histochem 2005;80:73-8. Torroni A, Schurr TG, Cabell MF, Brown MD, Neel JV, Larsen M, Smith DG, Vullo CM, Wallace DC. Asian affinities and continental radiation of the four founding native American mtdnas. Am J Hum Genet 1993;53: Underhill PA, Kivisild T. Use of Y chromosome and mitochondrial DNA population structure in tracing human migrations. Annu Rev Genet 2007;41: van Oven M, Kayser M. Updated phylogenetic tree of global human mitochondrial DNA variation. Hum Mutat 2009;30:E van Oven M. Revision of the mtdna tree and corresponding haplogroup nomenclature. Proc Natl Acad Sci 2010;107:E
100 Vigilant L, Stoneking M, Harpending H, Hawkes K, Wilson AC. African populations and the evolution of human mitochondrial DNA. Science 1991;253: Wakeley J. Substitution rate variation among sites in hypervariable region 1 of human mitochondrial DNA. J Mol Evol 1993;37: Wallace DC. Mitochondrial DNA variation in human evolution, degenerative disease, and aging. Am J Hum Genet 1995;57: Weissensteiner H, Pacher D, Kloss-Brandstätter A, Forer L, Specht G, Bandelt HJ, Kronenberg F, Salas A, Schönherr S. HaploGrep 2: mitochondrial haplogroup classification in the era of highthroughput sequencing. Nucleic Acids Res 2016a;44:W58-W63. Weissensteiner H, Forer L, Fuchsberger C, Schöpf B, Kloss- Brandstätter A, Specht G, Kronenberg F, Schönherr S. mtdna- Server: next-generation sequencing data analysis of human mitochondrial DNA in the cloud. Nucleic Acids Res 2016b;44:W64-9. Wills C. When did Eve live? An evolutionary detective story. Evolution 1995:49; Woischnik M, Moraes CT. Pattern of organization of human mitochondrial pseudogenes in the nuclear genome. Genome Res 2002;12; Yao YG, Kong QP, Bandelt HJ, Kivisild T, Zhang YP. Phylogeographic differentiation of mitochondrial DNA in Han Chinese. Am J Hum Genet 2002;70: Zhou Y, Guo F, Yu J, Liu F, Zhao J, Shen H, Zhao B, Jia F, Sun Z, Song H, Jiang X. Strategies for complete mitochondrial genome 89
101 sequencing on Ion Torrent PGM platform in forensic sciences. Forensic Sci Int Genet 2016;22:
102 Abstract Entire Mitochondrial DNA Sequencing on Next Generation Sequencing For the Korean Population Sohyung Park Department of Forensic Medicine The Graduate School Seoul National University Mitochondria DNA (mtdna) sequencing is an important analytic tool in understanding the genetic diversity that has been used for forensic identification as well as understanding the origin of humans in the phylogenetic study. Due to cost and time limitation, the traditional mtdna sequencing analysis was usually focused on the control region. As a result, the information of the genetic diversity had to be limited. In the wake of Human Genome Project, in order to handle the enormous amount of the genetic data, the need for a new analytic technology arose and the next generation sequencing was born. Thanks to this advanced technique, the complete mtdna sequencing could be available at a lower cost for less time, which allowed for an easier access to the information of genetic diversity - previously unavailable using the traditional method. Since then there have been many researches for mtdna genetic diversity in various people using this new analytic approach. And it is expected that it will strengthen the power of forensic identification based on 91
103 the genetic diversity on the complete mtdna sequencing. In terms of Koreans, most of the previous mtdna data were usually focused on the control region, such that the information of the genetic diversity in Korean was limited. Therefore, in keeping with research trends, it is necessary to study the genetic diversity in Korean based on complete mtdna sequences, using next generation sequencing. The purpose of this study is to understand the Korean genetic diversity of the mtdna by whole mtdna sequencing, and to confirm that it will strengthen the power of forensic identification in Koreans. 186 Koreans with common haplogroups in Korea based on the control region of mtdna were retreived, and whole mtdna sequencing using Ion Torrent Personal Genome Machine was performed on them. As a result, compared to people in East Asia including China, and other people in other continents, distribution of homoplasmic variants and point heteroplasmy of mtdna in Korean was generally similar to each other. Common variants between Korean and East Asia were similar, but they were different when compared to those of people in other continents. For haplogroup classification, and haplotypes determination, whole mtdna sequencing revealed better assignments compared to those based on the control region of mtdna, and some were reassigned into new haplogroups. These new assignments were also revealed similarlly in people in other countries. Given that the macrohaplogroup M (D, G, M) is major haplogroup in East Asians including Koreans, the complete mtdna sequencing is necessary to understand the genetic diversity, furthermore, the previous data focused on the control region might be incomplete. In addition, the result that the numbers of haplotypes were close to those of individuals after whole mtdna sequencing in 92
104 Korean, also contributed in strengthening the power of forensic identification. Also, the private variants in the same haplogroups and the fewer numbers of haplogroups than those of individuals, might suggest the possibility of some new potent subhaplogroups in Korean. Point heteroplasmy was also identified in Korean mtdna, and its distribution was similar to other studies. This study would be the driving force in understanding the genetic diversity of mtdna in Koreans in the future. Keywords: Forensic identification, Mitochondria, Mitochondrial DNA, Next generation sequencing, Korean Student Number :
105 표 [ 표 1] 186 명의한국인에서나타나는하플로그룹분포구성 하플로그룹 N 빈도 (%) 참고문헌 D4a 40 Allard et al. 2004; Lee et al. 2006; Jin et al D4a Lee et al D4b1 6 Lee et al D5b 3 Lee et al A5a Lee et al. 2006; Jin et al M7a Kivisild et al. 2002; Allard et al. 2004; Underhil et al. 2007; Jin et al M7b1 10 Lee et al F1b Lee et al G1a Tanaka et al. 2004; Lee et al B4a 16 Lee et al. 2006; Jin et al B4c1c 4 Lee et al N9a2a Tanaka et al. 2004; Lee et al M9;G;D Lee et al. 2006; Jin et al Y Lee et al. 2006; Jin et al. 2009; Hong et al total N, 각군의해당시료의숫자 94
106 [ 표 2] 186 명의한국인의인구학적특성 하플로그룹 N 지역정보 성별 지역모름남여모름 D4a 40 서울 2 제주 D4a3 3 제주 D4b1 6 제주 D5b A5a 20 제주 M7a1 9 제주 M7b1 10 제주 F1b1 16 제주 G1a1 19 서울 1 제주 B4a 16 제주 B4c1c 4 제주 N9a2a 11 부산 1 제주 M9;G;D Y1 12 제주 total N, 각군의해당시료의숫자 95
107 [ 표 3] 미토콘드리아 DNA 전체염기서열분석을위한길잡이 (Fendt L, et al., 2009) 미토콘드리아 DNA 전체염기서열을분석하기위하여, 미토콘드리아 DNA 염기서열을두부분으로나누고, 각부분은양끝부분일부가서로중복되게하여크기가약 8500 kb가되도록한다. 뉴클레오티드 2499번에서 10837번사이의염기서열부분을 A부분으로지정하고, 10672번에서 2669번사이의염기서열부분을 B부분으로지정한후, 각부분 5끝부분에서해당염기서열부위에상보적으로부합하는길잡이를제작하여 DNA 증폭을시행하였으며, 각길잡이서열은다음과같다. 이름염기서열 5 해당부분 PCR FampA AAATCTTACCCCGCCTGTTT A 부분 PCR RampA AATTAGGCTGTGGGTGGTTG A 부분 PCR FampB GCCATACTAGTCTTTGCCGC B 부분 PCR RampB GGCAGGTCAATTTCACTGGT B 부분 96
108 [ 표 4] 변이검출형식보고서 (VCF) 에서오검출된변이 변이검출 (Variant caller) 분석 해당변이가포함된 하플로그룹 변이의 전체수 보고안됨 71.1G B4a 3 247GA 247G 249del F1b1 16 보고안됨 455.1T D4 3 A5a, B4a, D4b, 513GCA 513G del F1b1, G1a1, 73 M7a1, M9;G;D 보고안됨 573.1C A5a, M9;G;D 3 보고안됨 960.1C N9a2a T 2150TA A A5a ATT AT,ATC 4248C A5a A 5894AC C M7a CACCCCCTCT 8270C del B4a, B4c C or 9540T/C 9540C D4 11 보고안됨 16182C, 16183C B4, F1b1, M7b, D5b 40 97
![[ 표 5] IGV 를이용한오검출된변이검토 변이검출 (Variant caller) 분석](/docs-images/92/109195273/images/109-0.jpg "IGV 로검토 보고안됨 71.")
109 [ 표 5] IGV 를이용한오검출된변이검토 변이검출 (Variant caller) 분석 IGV 로검토 보고안됨 71.1G (CE) 247GA 247G 249del (CE) 98
")

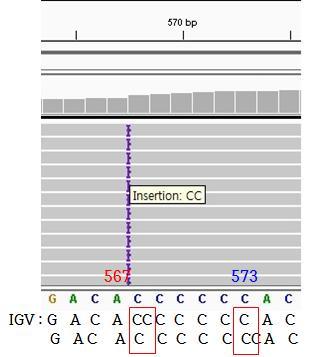
110 보고안됨 455.1T (CE) 513GCA 513G del 보고안됨 573.1C (CE) 99
")
")

111 2150T 2150TA A (Mt DB) 5894A 5894AC C (Mt DB) 8270CACCC CCTCT 8270C del 100
112 [ 표 6] 미토콘드리아 DNA 의조절부위와암호부위및전체에서나타난 동형세포질성변이 (homoplasmy variants) 의분포 하플로그룹 조절부위 암호부위 전체 미토콘드리아 DNA 암호부위 : 조절부위 N % N % N % 비율 D4a, D4a3, D4b1, D5b A5a M7a1, M7b F1b G1a B4a, B4c1c N9a2a M9;G;D Y Total N, 해당시료의수 101
113 [ 표 7] 하플로그룹에따른개별변이분포 하플로그룹 ( 전체염기서열 ) N 변이 D4a G D4b1a A, 4646C D4d C A5a A5a1a T F1b1a G G1a1a G G1a1a G G1a1a G G1a1a C, 9854C B4a T B G B4a1c1a T N9a2a C Y G N: 개체수 102
114 [ 표 8] 전체미토콘드리아 DNA 변이에따른하플로그룹세분화양상 조절부위 (N) D4a (40) 전체미토콘드리아 DNA (N) D4a (3), D4a1 (11), D4a1a1 (2), D4a1b (10), D4a1c (4), D4a2 (8), D4a2a (1), D4f1 (1) D4a3 (3) D4a3b (3) D4b1 (6) D4b1a1 (6) D5b (3) D5b1b (3) A5a (20) M7a1 (9) A5a (6), A5a1a (11), A5a1a1 (1), A5a1a2 (1), A5a3 (1) M7a1a (4), M7a1a1 (1), M7a1a5a (1), M7a1a9 (3) M7b1 (10) M7b1a1a (1), M7b1a1a1 (9) F1b1 (16) F1b1a (1), F1b1a1 (4), F1b1a1a (4), F1b1a1a1 (1), F1b1a1a2 (2), F1b1a2 (4) G1a1 (19) G1a1a (11), G1a1a1 (8) B4a (16) B4a (4), B (2), B4a1c1a (4), B4a1c1a1(6) B4c1c (4) B4c1c (4) N9a2a (11) N9a2a (10), N9a2a2 (1) M9;G;D (17) D4b2a2 (1), D4d (4), D4e2 (2), D4e2a (2), D4f1 (3), D4j (5) Y1 (12) Y1 (9), Y1a (1), Y1b1 (1), Y1b1a (1) Total (186) 186 N, 해당시료의수 103
115 [ 표 9] 하플로그룹을결정하는주요변이 하플로그룹 L L 미토콘드리아 DNA 주요변이 C146T C182T T4312C T10664C C10915T A11914G G13276A G16230A C152T A2758G C2885T G7146A T8468C L C195T A247G A825t T8655C A10688G C10810T G13105T T13506C G15301A A16129G T16187C C16189T L3 4 6 G4104A A7521G L3 4 T182C! T3594C T7256C T13650C T16278C L3 A769G A1018G C16311T M T489C C10400T T14783C G15043A M7 C6455T T9824C M7a T2626C C2772T T4385C A4958G G12771A, T16209C T16324 M7a1 M7a1a M7a1a XC G14364A T11017C A11084G T11722C M7a1a5a T13768C G7852A M7a1a9 G4048A M7b c M7b C4071T C12405T M7b1 A5351G G5460A T7684C G7853A G16129A! M7b1a1a T199C G4048A A4164G T6680C 104
116 T16297C C150T T12811C T16189C! M7b1a1a1 T10345C T16298C M9 M12 G G4491A T16362C G14569A G G709A A4833G T5108C T16362C G1 G1a G1a1 G1a1a G1a1a1 T8200C G15323A G15497A C150T C7867T A15860G T16325C A4793G G11914A! A827G M80 D D C4883T C5178a T16362C D4 G3010A C8414T C14668T D4a T152C! C3206T T8473C T14979C G16129A! D4a1 T10410C D4a1a1 G5261A G15314A D4a1b D4a1c D4a2 D4a2a D4a3 D4a3b D4b A13651G A7822G T12957C G3531A A8296G A10005G A5466G T16249C G7912A G8020A D4b1 C10181T T15440C A15951G G16319A D4b1a1 A14927G 105
117 T152C! A2246G T8450C A13827G A14091G G15217A A15805G D4b2 A1382c C8964T T9824a D4b2a2 A15524G G8251A C10104T D4d T3593C A5539G A6503c G7757A T9938C C12085T T13879C G16319A D4e D4e2 D4e2a D4f C11215T A15874G 573.XC A13716G T7270C D4f1 G2706A C4538T G8764A T11255C D4j G11696A D5 T16189C! C150T T1107C A5301G A10397G D5a b D5b A9180G C456T T681C C1048T A5153G A15724G D5b1b T6253C A3759G N G8701A C9540T G10398A C10873T A15301G! N9 G8417A N9a C150T G5231A A12358G N9a2 4 5 T16172C G12372A C16257a C16261T N9a2 N9a2a N9a2a2 T961C T15067C A16497G A14148G 106
118 Y G8392A A10398G! T14178C A14693G T16126C T16223C T16231C Y1 Y1a T146C! G3834A (C16266T) A7933G T16189C! Y1b1 A10097G C15460T G15221A Y1b1a C9278T A A235G A663G A1736G T4248C A4824G C8794T C16290T G16319A A5 A8563G C11536T A5a A G4655A C11647T C16187T! A5a1a C14944T G10801A A5a1a1 A5a1a2 A5a3 G5773A T12880C C8628T A12909G R R9 T12705C T16223C C3970T G13928c T16304C F A249d T6392 G10310A F1 G6962A T10609C G12406A C12882T F1b T16189C! T152C! C10976T C12633T G14476A C16232a T16249C T16311C! F1b1 F1b1a F1b1a1 F1b1a1a A4732G G5147A G16129A! C16344T A15954G C5049T 107
119 F1b1a1a1 F1b1a1a2 F1b1a2 T4705C A9389G C14629T R B T16189C! d B4 T16217C B C16261T B4a C16261T T5465C G9123A B4a1 B4a1c T10238C G709A B4a1c1a A9254G C15292T B4a1c1a1 B4c B4c1 G73c T89C T1119C G15346A C3497C B4c1c C150T T195C! A5441G A10398G! A13629G T15941C!: 계통발생학적으로조상의뉴클레오티드로 back mutation () 는개별하플로그룹내에서자주나타나는변이이지만주요변이로는아직불확실한변이이탈릭체로표시된변이는예비분류로추가적인염기서열정보로서개정될수있음. 삽입은. 으로, 결실은 d 로표시되었음. 밑줄 : 조절부위의변이 (Behar DM, et al., 2012, van Oven M, et al., 2009) 108
120 [ 표 10] 하플로타입의양상 Discriminating 고유한하플로타입수하플로그룹 (N) Capacity (%) 조절부위전체염기서열조절부위 전체염기서열 D4a (40) D4a3 (3) D4b1 (6) D5b (3) A5a (20) M7a1 (9) M7b1 (10) F1b1 (16) G1a1 (19) B4a (16) B4c1c (4) N9a2a (11) M9;G;D (17) Y1 (12) 전체 (186)
121 [ 표 11] 이형세포질성변이의분석 하플로그룹 부요소 Coverage 수준 (%) 위치 rcrs 주요소부요소 T T C 1 F1b1a1a A A G T T C 2 D4e2a T T C 3 D4a1b T T C G G A 4 D4a G G A T T C 5 N9a2a G G A 6 M7a1a5a T T C 7 B G G A 8 B4a C C A 9 G1a1a T T C A A G 10 A5a1a T C T 11 B4a1c1a T T C 12 D4a1b T T C 13 N9a2a T T C A G A 110
122 14 B4a1c1a A A G 15 D4f G G A 16 N9a2a A A G G G A 17 D4a1c C C T 18 D4a A A G 19 D4a T T C 20 D4j T T C T T C 21 D4j G G A 22 B4a1c1a C C T 23 D4a C C T 24 M7a1a A A G 25 N9a2a T T C 26 D4a2a G G A 27 D4j C C T 28 G1a1a G A G 29 A5a1a T T C 30 A5a1a C T C 31 M7b1a1a C C T 32 G1a1a C C T T T C 111
123 A A G C C T C T C 33 G1a1a C C T 34 M7a1a G A G 35 M7b1a1a A A G 36 D4b1a C C T 37 D4a1c C C T 38 B4a1c1a A G A 39 D4a3b C C T 40 A5a C T C Total 40 개체 52 곳의이형세포질성 rcrs: 개정캠프리지표준서열 (Revised Cambridge Reference Seqeunce) 112
124 [ 표 12] 점이형세포질성변이의분포 시료수 점이형세포질성변이의수 하플로그룹 전체 시료수 점이형세포질성시료수 ( 개체수 ) 조절 부위 암호 부위 전체 미토콘드리아 DNA D4a, D4a3, D4b1, D5b (21.15%) A5a 20 4 (20.00%) M7a1, M7b (26.32%) F1b (6.25%) G1a1 19 B4a, B4c1c 20 N9a2a 11 M9;G;D 17 4 (21.05%) 6 (30.00%) 4 (36.36%) 5 (29.41%) Y (0.00%) 전체 (21.51%) 16 (30.77%) 36 (69.23%) 52 (100%) 113
125 [ 표 13] 염기서열해독에서 coverage 비교 본연구, Seo 등의연구, Zhou 등의연구모두 Ion Torrent PGM을기반으로하는차세대염기서열분석기법을적용하여미토콘드리아염기서열을분석한연구로, coverage와미토콘드리아 DNA 염기서열에따른비교분석을하였다. 낮은 coverage를보이는부분에서공통적으로 NADH 탈수소효소아단위 5와아단위 6의유전자가위치하는암호부위가있음을알수있다. 본연구 Seo 등의연구 Zhou 등의연구 높은 coverage 낮은 coverage 평균 coverage 1,200X 810X 1,269X 114
126 [ 표 14] VCF 에서오검출된변이 : 본연구와다른연구와비교분석 변이본연구 Zhou 등 Parson 등 Seo 등 249del 247GA 247G 247GA 247G 513GCA 513GCA del 513G 513G C 보고안됨 567A 567ACCC, 567ACCCC, 567ACCCCC 573.1C, 573.2C 573.3C 4248C 4246TT AT/ATC 보고안됨 960.1C 보고안됨보고안됨 A 2150T 2150TA 2150T 2150TA C 5894A 5894AC 보고안됨 8270CACCCCCTCT 8289del 8270CACCCCCTC T 8270C 8270C 8286del del 8270CACCCCCTCTA C 8270CAC 16182C 보고안됨보고안됨보고안됨 16183C 보고안됨보고안됨보고안됨 115
127 [ 표 15] 동형세포질성변이의민족간비교 변이양상 본연구 ( 한국인 ) Zhou 등 ( 중국한족 ) Stoljarova 등 에스토니안인 King 등 ( 미국인 ) 연구대상 186 명 107 명 114 명 283 명 전체변이수 6,495 4,022 2,663 11,607 뉴클레오티드 위치수 455 곳 725 곳 512 곳 1,353 곳 1 개체당 평균변이수 조절부위 1, ,938 변이수 (25.9%) (22.18%) (22.8%) (25.3%) 암호부위 4,813 3,130 2,056 8,669 개 변이수 (74.1%) (77.82%) (77.2%) (74.7%) 치환변이 6,336 (97.55%) 3,919 (97.44%) 삽입결실 변이 (2.45%) (2.56%) 116
128 [ 표 16] 한국인및다른민족에서공통적으로나타나는변이비교 변이 본연구 ( 한국인 ) Zhou 등 ( 중국한족 ) Maruyama 등 ( 일본 ) Stoljarova 등 에스토니안인 King 등 ( 미국인 ) 73G 73G 73G 263G 263G 263G 263G 263G 750G 750G 750G 1438G 1438G 1438G 2706G 모두 4769G 4769G 4769G 4769G 7028G 7028G 8860G 8860G 11719G 14766T 11719G 14766T 15326G 15326G 15326G 15326G 489C 489C 16362C 73G G 2706G G 8701G 7028T C 9540C 11719A G 10398G 14766T 4747 절반 이상 10400T 10400T C 10873C T 12705T C 14783C A 15043A A 15301A T 16223T C 16519C 16519C 117
129 [ 표 17] 한국인과중국한족의하플로그룹세분화양상비교 본연구 ( 한국인 ) Zhou 등 ( 중국한족 ) 조절부위전체염기서열조절부위전체염기서열 A A5a A5a, A5a1a, A5a1a1, A5a1a2, A5a3 A A14 A15 A5a A5b, A A24 A15, A15a A5a B B4a B4c1c B4a, B , B4a1c1a, B4a1c1a1 B4c1c B4 B4b1 B4b1b c B4c1b B4c1b2a B4h1 B5b2a B5b2a1b B4d1a, B4d1 2 3, B4a3 B4b1a B4b1c B4c1b2c B4c1b2a B4h1 B5b2a B5b2a1b D D4e1 D4 D4j/D4 D4a1e D4b1a2a1 D4a5 D4a1e D4b2b D4a3b D D4a D4a3 D4b1 D4a, D4a1, D4a1a, D4a1b, D4a1c, D4a2, D4a2a, D4f1 D4a3b D4b1a1 D4b2b5 D4c D4c1b D4c1 D4g2a D4j3 D4k D4b1a2a1 D4b2b2 D4b2b5 D4c2 D4c1b D4 D4g2a1 D4o2a D4j3 D5a2a C4a1 D5a2a1 D5b D4o2a D5a2a, D5a2a1 D5c2 D5a2a1b M9;G;D D4b2a2, D4d, D4e2, D4e2a, D4f1, D4j M9a1a1c1b1 M74 D4j 118
130 F F1b1 F1b1a, F1b1a1, F1b1a1a, F1b1a1a1, F1b1a1a2, F1b1a2 G G1a1 G1a1, G1a1a1 M7a1 M7a1a, M7a1a1, M7a1a5a, M7a1a9 M M7b1a1a M7b1 M7b1a1a1 N N9a2a N9a2a, N9a2a2 F F1 F1a1 F1a1a F1b F1b1a F1b1b F1c1a1 F4a1a G1a1 G2a G2a1d G2a4 M10a1a1b M10a1a1b1 M45 M7b1a1 M7b1a1a1 M7c1 M8a M8a2 M8a2 3 M8a2a M8a2a1 M8a3a M9a1a1c M9a1a1c1b M9a1a2 M33 M9a b N9a N9a1 N9a1 3 F2, F2f F1g1 F1a1 F1a1a F1b1 F1b1a F1b1b F1c1a1 F4a1a G1a1 G2a2 G2a1d1a G2a4 M10a1a1b M10a1a1b1 M7c1a M7b1a1e1, M7b1a1h M7b1a1a1 M7c1c M8 M8a3 M8a3, M8a2 M8a3 M8a2a1 M8a3a1 M9a1a1c1a M9Ma1a M9a1a2 G2b1a, G1c G N9a9 N9a1 N9a3 119
131 [ 표 18] 하플로그룹재분류비교분석 하플로그룹 본연구 ( 한국인 ) Zhou 등 ( 중국한족 ) Stoljarova 등 ( 에스토니안인 ) King 등 ( 미국인 ) 새로운계통하플로그룹재분류같은계통다른하위하플로그룹재분류 M9;G;D à 4b2a2 M9;G;D à D4d M9;G;D à D4e2 M9;G;D à D4e2a M9;G;D à D4f1 M9;G;D à D4j M9a b à G U5b2a1a2 à H D4e1 à A2+64 M9a1a1c1b1 à D4 G3 à L2b1a4 M33 à 2b1a HV0 à V2 D4k à C4a1 M33c à A2+64 N2 à L2e1f N11a à L2a1a3 P5 à H32 C4b8 à C7 B4 à B2 B4 à B2a B4 à B2c1 B4 à B2f B4c1a2a à B2b H2a2a à H3 H2a2a à H1bk H2a2a à H56a1 H2a2a1d àh1c7 H3v à H76 K1c à K2b I2 3 à I2 120
132 [ 표 19] 점이형세포질성 (PHP) 에대한비교 본연구 Zhou 등 2016 Li 등 2014 King 등 2014 Li 등 2015 Skonieczna 등 2015 Sosa 등 2012 실험 방법 NGS/ PGM NGS/ PGM NGS/ Illumina GA II NGS/ Illumina MiSeq NGS/ Illumina HiSeq NGS/ Roche 454 NGS/ Roche 454 기준 mtdnaserver >40X no numt 15% 10-20% 18% >7X >0.5% allele frequency >50X 2% 10% 관찰 범위 45.93% 8-46% 2-43% 9-50% >40X 67X± coverage 46-36,444X >65X 개체수 PHP 개체 (21.51%) (20.56%) (24.24%) (24.03%) (22.30%) (32.00%) 26 (65.00%) PHP 전체 PHP 조절부위 PHP 암호부위 (30.77%) (25.00%) (35.14%) (34.83%) (37.14%) (56.52%) (69.23%) (75.00%) (64.86%) (65.17%) (62.86%) (43.48%) 13 (18.31%) 58 (81.69%) 최대 PHP / 개체
133 그림 [ 그림 1] 미토콘드리아 DNA 의구조 ( 미토콘드리아 DNA는무거운가닥과가벼운가닥으로구성된 2중원형의구조로되어있으며, D-Loop 부위등복제와전사가시작되는부위가있는조절부위와유전자가암호화되어있는암호부위로구성되어있다. 122
![[ 그림 2] 미토콘드리아 DNA 에따른인류의이동과분포 (www.mitomap.](/docs-images/92/109195273/images/134-0.jpg "org) 인류는아프리카에서이주하기시작하여중동지역을거쳐일부는유럽중부와북부로, 일부는아시아남쪽과동남쪽으로이동하였으며, 일부는중국지역에서인도네시아섬지역을거쳐뉴기니아, 호주로이주하였고,")
134 [ 그림 2] 미토콘드리아 DNA 에따른인류의이동과분포 ( 인류는아프리카에서이주하기시작하여중동지역을거쳐일부는유럽중부와북부로, 일부는아시아남쪽과동남쪽으로이동하였으며, 일부는중국지역에서인도네시아섬지역을거쳐뉴기니아, 호주로이주하였고, 아시아동쪽끝을따라북쪽으로베링해협을건너알래스카에서남쪽으로진행하여북미, 중미, 남미로이주하여최종뉴기니아와오스트레일리아북쪽주면의섬에서북태평양 ( 폴리네시아, 하와이등 ) 으로이동하였을것으로본다. 123
![[ 그림 3] 인류계통발생나무 (www.mitomap.](/docs-images/92/109195273/images/135-0.jpg "org) 여러연구를통해얻어진미토콘드리아 DNA의염기서열분석결과를토대로세계의각지역에따라분류되는하플로그룹과그계통발생학적나무가수립되었다.")
135 [ 그림 3] 인류계통발생나무 ( 여러연구를통해얻어진미토콘드리아 DNA의염기서열분석결과를토대로세계의각지역에따라분류되는하플로그룹과그계통발생학적나무가수립되었다. 계통발생학적뿌리는아프리카에두었으며여기서부터아시아와유럽등지로분지되는것으로보았다. 아시아에서는하플로그룹 M에서각세부군으로분류되는것을알수있다. 124
136 [ 그림 4] 한국인에서알려진하플로그룹의빈도 (Lee HY, et al., 2006) 593명의한국인을대상으로각미토콘드리아 DNA 조절부위의염기서열을분석한결과에의거한것으로이를통해한국인에서주로나타나는것으로알려진각하플로그룹별빈도를보여주고있다. 가장흔한하플로그룹으로 D4, D4a, N9a2 등이있었다. 125
![[ 그림 5] 동아시아인에서주요하플로그룹의빈도 (Allard MW, et al.](/docs-images/92/109195273/images/137-0.jpg ", 2004) 동아시아인구 ( 중국인 356명, 한국인 182명, 일본인 163명 ) 를대상으로시행한미토콘드리아 DNA 조절부위염기서열분석결과에의거한것으로중국, 한국일본인구에서주로나타나는하플로그룹의빈도를보여주고있다.")
137 [ 그림 5] 동아시아인에서주요하플로그룹의빈도 (Allard MW, et al., 2004) 동아시아인구 ( 중국인 356명, 한국인 182명, 일본인 163명 ) 를대상으로시행한미토콘드리아 DNA 조절부위염기서열분석결과에의거한것으로중국, 한국일본인구에서주로나타나는하플로그룹의빈도를보여주고있다. ( 검정색막대 : 중국, 사선막대 : 한국, 흰색막대 : 일본 ) 한국인에서가장많았던하플로그룹은 D4였으며, 이외하플로그룹 A, D4a, D5 등이있었다. 126
![[ 그림 6] Ion Torrent PGM (Personal Genome Machine) 기법 차세대염기서열분석기법으로 Ion](/docs-images/92/109195273/images/138-0.jpg "Torrent Personal Genome Machine (PGM) 을이용하여초병렬염기서열분석을시행하였으며그방법은 다음과같았다.")
138 [ 그림 6] Ion Torrent PGM (Personal Genome Machine) 기법 차세대염기서열분석기법으로 Ion Torrent Personal Genome Machine (PGM) 을이용하여초병렬염기서열분석을시행하였으며그방법은 다음과같았다. 127
법학박사학위논문 실손의료보험연구 2018 년 8 월 서울대학교대학원 법과대학보험법전공 박성민
 저작자표시 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
행정학석사학위논문 공공기관기관장의전문성이 조직의성과에미치는영향 년 월 서울대학교행정대학원 행정학과행정학전공 유진아
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
문학석사학위논문 존밀링턴싱과이효석의 세계주의비교 로컬 을중심으로 년 월 서울대학교대학원 협동과정비교문학 이유경
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
경영학석사학위논문 투자발전경로이론의가설검증 - 한국사례의패널데이타분석 년 8 월 서울대학교대학원 경영학과국제경영학전공 김주형
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물
 저작자표시 - 비영리 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 - 비영리 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
i
 저작자표시 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
Precipitation prediction of numerical analysis for Mg-Al alloys
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
 저작자표시 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비
 저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는,
저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니
 저작자표시 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
행정학박사학위논문 목표모호성과조직행태 - 조직몰입, 직무만족, 공직봉사동기에미치는 영향을중심으로 - 년 월 서울대학교대학원 행정학과행정학전공 송성화
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원
 저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는, 이저작물과동일한이용허락조건하에서만배포할수있습니다.
저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는, 이저작물과동일한이용허락조건하에서만배포할수있습니다.
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
교육학석사학위논문 윤리적입장에따른학교상담자의 비밀보장예외판단차이분석 년 월 서울대학교대학원 교육학과교육상담전공 구승영
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비
 저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는,
저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 변경금지. 귀
 저작자표시 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우, 이저작물에적용된이용허락조건을명확하게나타내어야합니다.
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
2016 학년도약학대학면접문제해설 문제 2 아래의질문에 3-4분이내로답하시오. 표피성장인자수용체 (epidermal growth factor receptor, EGFR) 는수용체티로신인산화효소군 (receptor tyrosine kinases, RTKs) 의일종으로서세
 는수용체티로신인산화효소군 (receptor tyrosine kinases, RTKs) 의일종으로서세") 본문제에대한지적소유권은동국대학교에있습니다. 본교의서면허락없이무단으로출판, 게재, 사용할수없습니다. 문제 2 2016 학년도약학대학면접문제 아래의질문에 3-4 분이내로답하시오. 표피성장인자수용체 (epidermal growth factor receptor, EGFR) 는수용체티로신 인산화효소군 (receptor tyrosine kinases, RTKs) 의일종으로서세포의생존과증식
본문제에대한지적소유권은동국대학교에있습니다. 본교의서면허락없이무단으로출판, 게재, 사용할수없습니다. 문제 2 2016 학년도약학대학면접문제 아래의질문에 3-4 분이내로답하시오. 표피성장인자수용체 (epidermal growth factor receptor, EGFR) 는수용체티로신 인산화효소군 (receptor tyrosine kinases, RTKs) 의일종으로서세포의생존과증식
농학석사학위논문 폴리페닐렌설파이드복합재료의기계적및열적 특성에영향을미치는유리섬유 환원된 그래핀옥사이드복합보강재에관한연구 The combined effect of glass fiber/reduced graphene oxide reinforcement on the mecha
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원
 저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는, 이저작물과동일한이용허락조건하에서만배포할수있습니다.
저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는, 이저작물과동일한이용허락조건하에서만배포할수있습니다.
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
 저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는,
저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는,
 Ⅰ. 석면 1 1) American Geological Institute, Glossary of geology, 2008, http://glossary.agiweb.org 2) US OSHA standard 29CFR1910.1001(b) 2 석면분석전문가양성교육교재 : 편광현미경을이용한고형시료중석면분석 1) Cornelis Klein, The Manual
Ⅰ. 석면 1 1) American Geological Institute, Glossary of geology, 2008, http://glossary.agiweb.org 2) US OSHA standard 29CFR1910.1001(b) 2 석면분석전문가양성교육교재 : 편광현미경을이용한고형시료중석면분석 1) Cornelis Klein, The Manual
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
<4D F736F F F696E74202D20332DB5B9BFACBAAFC0CC2DB0A8BCF6BAD0BFAD2DBFB0BBF6C3BCC0CCBBF3205BC8A3C8AF20B8F0B5E55D>
 주제 3. 유전자돌연변이, 감수분열, 염색체이상 생명과학 10 장중간부분 생명과학 8 장뒷부분 유전체의 DNA 염기서열해독이끝난생물체와유전체크기, 유전자수 1 1. 돌연변이에의해유전정보가달라질수있다 돌연변이 (mutation) 란 DNA 의염기서열에변화가일어난것이다 돌연변이는 염색체상의넓은부위나 한개의뉴클레오티드에만생길수도있다 사람의경우, 생식세포 ( 정자,
주제 3. 유전자돌연변이, 감수분열, 염색체이상 생명과학 10 장중간부분 생명과학 8 장뒷부분 유전체의 DNA 염기서열해독이끝난생물체와유전체크기, 유전자수 1 1. 돌연변이에의해유전정보가달라질수있다 돌연변이 (mutation) 란 DNA 의염기서열에변화가일어난것이다 돌연변이는 염색체상의넓은부위나 한개의뉴클레오티드에만생길수도있다 사람의경우, 생식세포 ( 정자,
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
<BBFDC8ADC7D02E687770>
 대상본과 1 학년담당교수손정원 2014. 4. 1. 3/4 교시장소 1 의학관제 1 강의실 2. 수업주제 단백질 folding, targeting, 분해 1) 단백질 folding의기본동력에대해서이해한다. 2) chaperone의종류와작용기전에대하여설명한다. 3) proteasome의구조와 proteasome에의한단백질분해에대하여설명한다. 4) ER targeting
대상본과 1 학년담당교수손정원 2014. 4. 1. 3/4 교시장소 1 의학관제 1 강의실 2. 수업주제 단백질 folding, targeting, 분해 1) 단백질 folding의기본동력에대해서이해한다. 2) chaperone의종류와작용기전에대하여설명한다. 3) proteasome의구조와 proteasome에의한단백질분해에대하여설명한다. 4) ER targeting
 유전자의전승 (Genetic Heritage) 일러두기 1 서론 3 제 장더러브렛유전자풀 - 기원 9 제 장더러브렛종을형성한위대한선대마들 59 제 장스피드와스태미나인자 95 제 장심장의크기와 X 요인의유전가능성 115 제 장부계혈통의생존 - 생식력과주기적기능 139 제 장세포의미토콘드리아와모계의중요성 175 제 장 BLANDFORD 가스태미나에미친영향
유전자의전승 (Genetic Heritage) 일러두기 1 서론 3 제 장더러브렛유전자풀 - 기원 9 제 장더러브렛종을형성한위대한선대마들 59 제 장스피드와스태미나인자 95 제 장심장의크기와 X 요인의유전가능성 115 제 장부계혈통의생존 - 생식력과주기적기능 139 제 장세포의미토콘드리아와모계의중요성 175 제 장 BLANDFORD 가스태미나에미친영향
행정학석사학위논문 외국인주민생활만족도의 영향요인연구 년 월 서울대학교대학원 행정학과행정학전공 최은영
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
Chapter 26
 11 주 RNA 합성 11.1 DNA-dependent synthesis of RNA: Bacteria에서의 transcription l RNA polymerase (5 subunits로구성 : 2α, β, β, σ) l 효소에의한 RNA 합성의특징 - Complementary sequence to template DNA - RNA chain의합성방향 : 5
11 주 RNA 합성 11.1 DNA-dependent synthesis of RNA: Bacteria에서의 transcription l RNA polymerase (5 subunits로구성 : 2α, β, β, σ) l 효소에의한 RNA 합성의특징 - Complementary sequence to template DNA - RNA chain의합성방향 : 5
저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비
 저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는,
저작자표시 - 비영리 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는,
PowerPoint 프레젠테이션
 mrna (messenger RNA) : 양이가장적다 ( 전체 RNA 중 5-10% 이하 ) : mrna의염기서열은단백질의아미노산순서를지정 : 성장하는세포는수많은다른단백질을필요로함으로 mrna가만들어져단백질이합성되게한후 nucleotide가재생될수있도록분해된다 (trna, rrna도재생 ) : 5 말단의끝이 7-methyl guanosine triphosphate
mrna (messenger RNA) : 양이가장적다 ( 전체 RNA 중 5-10% 이하 ) : mrna의염기서열은단백질의아미노산순서를지정 : 성장하는세포는수많은다른단백질을필요로함으로 mrna가만들어져단백질이합성되게한후 nucleotide가재생될수있도록분해된다 (trna, rrna도재생 ) : 5 말단의끝이 7-methyl guanosine triphosphate
<4D F736F F F696E74202D2035BBF3C6F2C7FC5FBCF8BCF6B9B0C1FA2E BC8A3C8AF20B8F0B5E55D>
 5. 상평형 : 순수물질 이광남 5. 상평형 : 순수물질 상전이 phase transition 서론 ~ 조성의변화없는상변화 5. 상평형 : 순수물질 전이열역학 5. 안정성조건 G ng ng n G G 자발적변화 G < 0 G > G or 물질은가장낮은몰Gibbs 에너지를갖는상 가장안정한상 으로변화하려는경향 5. 상평형 : 순수물질 3 5. 압력에따른Gibbs
5. 상평형 : 순수물질 이광남 5. 상평형 : 순수물질 상전이 phase transition 서론 ~ 조성의변화없는상변화 5. 상평형 : 순수물질 전이열역학 5. 안정성조건 G ng ng n G G 자발적변화 G < 0 G > G or 물질은가장낮은몰Gibbs 에너지를갖는상 가장안정한상 으로변화하려는경향 5. 상평형 : 순수물질 3 5. 압력에따른Gibbs
치의학석사학위논문 치의학대학원학생의장애환자에 대한인식조사 년 월 서울대학교치의학대학원 치의학과 박상억
 저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는, 이저작물과동일한이용허락조건하에서만배포할수있습니다.
저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는, 이저작물과동일한이용허락조건하에서만배포할수있습니다.
<5BB0EDB3ADB5B55D32303131B3E2B4EBBAF12DB0ED312D312DC1DFB0A32DC0B6C7D5B0FAC7D02D28312E28322920BAF2B9F0B0FA20BFF8C0DAC0C720C7FCBCBA2D3031292D3135B9AEC7D72E687770>
 고1 융합 과학 2011년도 1학기 중간고사 대비 다음 글을 읽고 물음에 답하시오. 1 빅뱅 우주론에서 수소와 헬륨 의 형성에 대한 설명으로 옳은 것을 보기에서 모두 고른 것은? 4 서술형 다음 그림은 수소와 헬륨의 동위 원 소의 을 모형으로 나타낸 것이. 우주에서 생성된 수소와 헬륨 의 질량비 는 약 3:1 이. (+)전하를 띠는 양성자와 전기적 중성인 중성자
고1 융합 과학 2011년도 1학기 중간고사 대비 다음 글을 읽고 물음에 답하시오. 1 빅뱅 우주론에서 수소와 헬륨 의 형성에 대한 설명으로 옳은 것을 보기에서 모두 고른 것은? 4 서술형 다음 그림은 수소와 헬륨의 동위 원 소의 을 모형으로 나타낸 것이. 우주에서 생성된 수소와 헬륨 의 질량비 는 약 3:1 이. (+)전하를 띠는 양성자와 전기적 중성인 중성자
º´¹«Ã»Ã¥-»ç³ªÀÌ·Î
 솔직히 입대하기 전까지만 해도 왜 그렇게까지 군대를 가려고하냐, 미친 것 아니냐는 소리도 많이 들었다. 하지만 나는 지금 그 때의 선택을 후회하지 않는다. 내가 선택한 길이기에 후회는 없다. 그런 말을 하던 사람들조차 지금의 내 모습을 보고 엄지 손가락을 치켜세운다. 군대는 하루하루를 소종하게 생각 할 수 있게 만들어 주었고, 점점 변해가는 내 모습을 보며
솔직히 입대하기 전까지만 해도 왜 그렇게까지 군대를 가려고하냐, 미친 것 아니냐는 소리도 많이 들었다. 하지만 나는 지금 그 때의 선택을 후회하지 않는다. 내가 선택한 길이기에 후회는 없다. 그런 말을 하던 사람들조차 지금의 내 모습을 보고 엄지 손가락을 치켜세운다. 군대는 하루하루를 소종하게 생각 할 수 있게 만들어 주었고, 점점 변해가는 내 모습을 보며
<4D6963726F736F667420506F776572506F696E74202D204F435720B5EEB7CFB0ADC0C7C0DAB7E12DBBFDC8B0BCD3C0C720BBFDB8EDB0FAC7D0>
 생활 속의 생명과학 (Biological Science for Life) 13강. DNA 검사 13 주차 강의 DNA 검사 - 법의학 과학 앞에 완전범죄는 더 이상 없다 [아시아경제] 2009.4.15 범인라고 확신하고 추적하던 용의자의 DNA가 현장에서 발견된 것과 일치하지 않는다면, 형사는 용의자를 더 이상 잡아둘 수 없다. 결정적인 증거가 없기 때문이다.
생활 속의 생명과학 (Biological Science for Life) 13강. DNA 검사 13 주차 강의 DNA 검사 - 법의학 과학 앞에 완전범죄는 더 이상 없다 [아시아경제] 2009.4.15 범인라고 확신하고 추적하던 용의자의 DNA가 현장에서 발견된 것과 일치하지 않는다면, 형사는 용의자를 더 이상 잡아둘 수 없다. 결정적인 증거가 없기 때문이다.
 저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는, 이저작물과동일한이용허락조건하에서만배포할수있습니다.
저작자표시 - 동일조건변경허락 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 이차적저작물을작성할수있습니다. 이저작물을영리목적으로이용할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 동일조건변경허락. 귀하가이저작물을개작, 변형또는가공했을경우에는, 이저작물과동일한이용허락조건하에서만배포할수있습니다.
Journal of Life Science 2011, Vol. 21. No μ μ
 Journal of Life Science 2011 Vol. 21. No. 8. 1120~1126 ISSN : 1225-9918 DOI : http://dx.doi.org/10.5352/jls.2011.21.8.1120 μ μ μ α β Journal of Life Science 2011, Vol. 21. No. 8 1121 μ μ 1122 생명과학회지 2011,
Journal of Life Science 2011 Vol. 21. No. 8. 1120~1126 ISSN : 1225-9918 DOI : http://dx.doi.org/10.5352/jls.2011.21.8.1120 μ μ μ α β Journal of Life Science 2011, Vol. 21. No. 8 1121 μ μ 1122 생명과학회지 2011,
농림축산식품부장관귀하 본보고서를 미생물을활용한친환경작물보호제및비료의제형화와현장적용매뉴 얼개발 ( 개발기간 : ~ ) 과제의최종보고서로제출합니다 주관연구기관명 : 고려바이오주식회사 ( 대표자 ) 김영권 (
 과제의최종보고서로제출합니다 주관연구기관명 : 고려바이오주식회사 ( 대표자 ) 김영권 (") 농림축산식품부장관귀하 본보고서를 미생물을활용한친환경작물보호제및비료의제형화와현장적용매뉴 얼개발 ( 개발기간 :2014. 7. 29 ~ 2016. 7. 28.) 과제의최종보고서로제출합니다. 2016. 7. 28. 주관연구기관명 : 고려바이오주식회사 ( 대표자 ) 김영권 ( 인 ) 협동연구기관명 : 목원대학교산학협력단 ( 대표자 ) 고대식 ( 인 ) 협동연구기관명
농림축산식품부장관귀하 본보고서를 미생물을활용한친환경작물보호제및비료의제형화와현장적용매뉴 얼개발 ( 개발기간 :2014. 7. 29 ~ 2016. 7. 28.) 과제의최종보고서로제출합니다. 2016. 7. 28. 주관연구기관명 : 고려바이오주식회사 ( 대표자 ) 김영권 ( 인 ) 협동연구기관명 : 목원대학교산학협력단 ( 대표자 ) 고대식 ( 인 ) 협동연구기관명
Sequences with Low Correlation
 레일리페이딩채널에서의 DPC 부호의성능분석 * 김준성, * 신민호, * 송홍엽 00 년 7 월 1 일 * 연세대학교전기전자공학과부호및정보이론연구실 발표순서 서론 복호화방법 R-BP 알고리즘 UMP-BP 알고리즘 Normalied-BP 알고리즘 무상관레일리페이딩채널에서의표준화인수 모의실험결과및고찰 결론 Codig ad Iformatio Theory ab /15
레일리페이딩채널에서의 DPC 부호의성능분석 * 김준성, * 신민호, * 송홍엽 00 년 7 월 1 일 * 연세대학교전기전자공학과부호및정보이론연구실 발표순서 서론 복호화방법 R-BP 알고리즘 UMP-BP 알고리즘 Normalied-BP 알고리즘 무상관레일리페이딩채널에서의표준화인수 모의실험결과및고찰 결론 Codig ad Iformatio Theory ab /15
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
제 53 회서울특별시과학전람회 예선대회작품설명서 본선대회작품설명서 쓰나미의피해를최소화시키는건물과 건물배치에대한탐구 출품번호 S-504 출품분야학생부출품부문지구과학 학교명학년 ( 직위 ) 성명
 성명") 제 53 회서울특별시과학전람회 예선대회작품설명서 본선대회작품설명서 쓰나미의피해를최소화시키는건물과 건물배치에대한탐구 출품번호 S-504 출품분야학생부출품부문지구과학 2012. 5. 14. 학교명학년 ( 직위 ) 성명 - 1 - 그림 1 쓰나미의발생과정 그림 2 실제쓰나미의사진 ρ - 2 - 그림 3 땅을파는모습그림 4 완성된수조의모습 - 3 - 그림 5 삼각기둥그림
제 53 회서울특별시과학전람회 예선대회작품설명서 본선대회작품설명서 쓰나미의피해를최소화시키는건물과 건물배치에대한탐구 출품번호 S-504 출품분야학생부출품부문지구과학 2012. 5. 14. 학교명학년 ( 직위 ) 성명 - 1 - 그림 1 쓰나미의발생과정 그림 2 실제쓰나미의사진 ρ - 2 - 그림 3 땅을파는모습그림 4 완성된수조의모습 - 3 - 그림 5 삼각기둥그림
세계 비지니스 정보
 - i - ii - iii - iv - v - vi - vii - viii - ix - 1 - 2 - 3 - - - - - - - - - - 4 - - - - - - 5 - - - - - - - - - - - 6 - - - - - - - - - 7 - - - - 8 - 9 - 10 - - - - - - - - - - - - 11 - - - 12 - 13 -
- i - ii - iii - iv - v - vi - vii - viii - ix - 1 - 2 - 3 - - - - - - - - - - 4 - - - - - - 5 - - - - - - - - - - - 6 - - - - - - - - - 7 - - - - 8 - 9 - 10 - - - - - - - - - - - - 11 - - - 12 - 13 -
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
Microsoft PowerPoint - 26.pptx
 이산수학 () 관계와그특성 (Relations and Its Properties) 2011년봄학기 강원대학교컴퓨터과학전공문양세 Binary Relations ( 이진관계 ) Let A, B be any two sets. A binary relation R from A to B, written R:A B, is a subset of A B. (A 에서 B 로의이진관계
이산수학 () 관계와그특성 (Relations and Its Properties) 2011년봄학기 강원대학교컴퓨터과학전공문양세 Binary Relations ( 이진관계 ) Let A, B be any two sets. A binary relation R from A to B, written R:A B, is a subset of A B. (A 에서 B 로의이진관계
통신1310_01-도비라및목차1~9
 ISSN 5-693 13. 1 13. 1 3 1 3 1 i .75 1.5.75 1. 1..9.9 7.5 ) 7.5 ) 3. 1.5 1.5 9. ) 1. ) ii 8 6 8 6 - - 3 1 1 11 1 9 8 7 iii 6 5 6 5 3 3 1 8 1 8 1 1 6 6-1 -1 - - iv . 1.5 1.. 1.5 1..5. -.5.5. -.5
ISSN 5-693 13. 1 13. 1 3 1 3 1 i .75 1.5.75 1. 1..9.9 7.5 ) 7.5 ) 3. 1.5 1.5 9. ) 1. ) ii 8 6 8 6 - - 3 1 1 11 1 9 8 7 iii 6 5 6 5 3 3 1 8 1 8 1 1 6 6-1 -1 - - iv . 1.5 1.. 1.5 1..5. -.5.5. -.5
Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정
![Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정](/thumbs/88/116783310.jpg "Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정") Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정보를 RNA로옮겨주는과정 ex) retrovirus 2. DNA Replication ( 유전자복제
Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정보를 RNA로옮겨주는과정 ex) retrovirus 2. DNA Replication ( 유전자복제
<4D F736F F F696E74202D20322DC0AFC0FCC0DAB9DFC7F6C0C7C1B6C0FD205BC8A3C8AF20B8F0B5E55D>
 주제 2. 유전자발현의조절 생명과학 11 장앞부분 Figure 06.01: The minimum gene number required for any type of organism increases with its complexity. 1 인간게놈 DNA 중 : 유전자 ( 엑손 + 인트론 ): 25% 엑손 : 1%, 인트론 : 24% 나머지는유전자가아닌 DNA
주제 2. 유전자발현의조절 생명과학 11 장앞부분 Figure 06.01: The minimum gene number required for any type of organism increases with its complexity. 1 인간게놈 DNA 중 : 유전자 ( 엑손 + 인트론 ): 25% 엑손 : 1%, 인트론 : 24% 나머지는유전자가아닌 DNA
°ø±â¾Ð±â±â
 20, 30, 40 20, 30, 40 1 2 3 4 5 6 7 8 9 10 3.1 6.3 9.4 12.6 15.7 18.8 22.0 25.1 28.3 31.4 2.4 4.7 7.1 9.4 11.8 14.1 16.5 18.8 21.2 23.6 7.1 14.1 21.2 28.3 35.3 42.4 49.5 56.5 63.6 70.7 5.9 11.9 17.8 23.7
20, 30, 40 20, 30, 40 1 2 3 4 5 6 7 8 9 10 3.1 6.3 9.4 12.6 15.7 18.8 22.0 25.1 28.3 31.4 2.4 4.7 7.1 9.4 11.8 14.1 16.5 18.8 21.2 23.6 7.1 14.1 21.2 28.3 35.3 42.4 49.5 56.5 63.6 70.7 5.9 11.9 17.8 23.7
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
미생물분류는형태적특징, 생리 생화학적성질과상태, 화학분류학적성질과상태등을이용하여구분하는것이일반적이지만, 이와같은방법을이용하면많은시간을필요로한다. 또한분류가힘든경우나, 정확하지못한결과를얻는경우도있다. 최근미생물분류에도분자생물학적인방법을이용하여, 미생물이가지고있는 DNA를
 Code RR180A - 연구용 - Bacterial 16S rdna PCR Kit 사용설명서 v201011da 미생물분류는형태적특징, 생리 생화학적성질과상태, 화학분류학적성질과상태등을이용하여구분하는것이일반적이지만, 이와같은방법을이용하면많은시간을필요로한다. 또한분류가힘든경우나, 정확하지못한결과를얻는경우도있다. 최근미생물분류에도분자생물학적인방법을이용하여, 미생물이가지고있는
Code RR180A - 연구용 - Bacterial 16S rdna PCR Kit 사용설명서 v201011da 미생물분류는형태적특징, 생리 생화학적성질과상태, 화학분류학적성질과상태등을이용하여구분하는것이일반적이지만, 이와같은방법을이용하면많은시간을필요로한다. 또한분류가힘든경우나, 정확하지못한결과를얻는경우도있다. 최근미생물분류에도분자생물학적인방법을이용하여, 미생물이가지고있는
< C0DAC0B2C5BDB1B820BFEEBFB520B8DEB4BABEF32D33C2F720C6EDC1FD2E687770>
 과학영재의창의적탐구능력배양을위한 R&E 프로그램기획 운영핸드북 Handbook of Annual Planning and Implementing R&E Program for the Talented 2017 과학영재창의연구 (R&E) 지원센터 이핸드북은과학고와과학영재학교의연간 R&E 프로그램기획 운영을효과적으로지원하기위해개발된것으로, 한국과학창의재단지정과학영재창의연구
과학영재의창의적탐구능력배양을위한 R&E 프로그램기획 운영핸드북 Handbook of Annual Planning and Implementing R&E Program for the Talented 2017 과학영재창의연구 (R&E) 지원센터 이핸드북은과학고와과학영재학교의연간 R&E 프로그램기획 운영을효과적으로지원하기위해개발된것으로, 한국과학창의재단지정과학영재창의연구
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
교육학석사학위논문 중학교자유학기제를위한 음악과산업 수업설계 및지도방법에대한연구 년 월 서울대학교대학원 협동과정음악교육전공 염우정
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
동아시아국가들의실질환율, 순수출및 경제성장간의상호관계비교연구 : 시계열및패널자료인과관계분석
 동아시아국가들의실질환율, 순수출및 경제성장간의상호관계비교연구 : 시계열및패널자료인과관계분석 목차 I. 서론 II. 동아시아각국의무역수지, 실질실효환율및 GDP간의관계 III. 패널데이터를이용한 Granger인과관계분석 IV. 개별국실증분석모형및 TYDL을이용한 Granger 인과관계분석 V. 결론 참고문헌 I. 서론 - 1 - - 2 - - 3 - - 4
동아시아국가들의실질환율, 순수출및 경제성장간의상호관계비교연구 : 시계열및패널자료인과관계분석 목차 I. 서론 II. 동아시아각국의무역수지, 실질실효환율및 GDP간의관계 III. 패널데이터를이용한 Granger인과관계분석 IV. 개별국실증분석모형및 TYDL을이용한 Granger 인과관계분석 V. 결론 참고문헌 I. 서론 - 1 - - 2 - - 3 - - 4
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할
 저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
저작자표시 - 비영리 - 변경금지 2.0 대한민국 이용자는아래의조건을따르는경우에한하여자유롭게 이저작물을복제, 배포, 전송, 전시, 공연및방송할수있습니다. 다음과같은조건을따라야합니다 : 저작자표시. 귀하는원저작자를표시하여야합니다. 비영리. 귀하는이저작물을영리목적으로이용할수없습니다. 변경금지. 귀하는이저작물을개작, 변형또는가공할수없습니다. 귀하는, 이저작물의재이용이나배포의경우,
Jkbcs016(92-97).hwp
.hwp") Expression of bcl-2 and Apoptosis and Its Relationship to Clinicopathological Prognostic Factors in Breast Cancer - A Study with Long Term Follow-up correlated with the survival rate.(journal of Korean
Expression of bcl-2 and Apoptosis and Its Relationship to Clinicopathological Prognostic Factors in Breast Cancer - A Study with Long Term Follow-up correlated with the survival rate.(journal of Korean
제 3강 역함수의 미분과 로피탈의 정리
 제 3 강역함수의미분과로피탈의정리 역함수의미분 : 두실수 a b 와폐구갂 [ ab, ] 에서 -이고연속인함수 f 가 ( a, b) 미분가능하다고가정하자. 만일 f '( ) 0 이면역함수 f 은실수 f( ) 에서미분가능하고 ( f )'( f ( )) 이다. f '( ) 에서 증명 : 폐구갂 [ ab, ] 에서 -이고연속인함수 f 는증가함수이거나감소함수이다 (
제 3 강역함수의미분과로피탈의정리 역함수의미분 : 두실수 a b 와폐구갂 [ ab, ] 에서 -이고연속인함수 f 가 ( a, b) 미분가능하다고가정하자. 만일 f '( ) 0 이면역함수 f 은실수 f( ) 에서미분가능하고 ( f )'( f ( )) 이다. f '( ) 에서 증명 : 폐구갂 [ ab, ] 에서 -이고연속인함수 f 는증가함수이거나감소함수이다 (
Microsoft PowerPoint Relations.pptx
 이산수학 () 관계와그특성 (Relations and Its Properties) 2010년봄학기강원대학교컴퓨터과학전공문양세 Binary Relations ( 이진관계 ) Let A, B be any two sets. A binary relation R from A to B, written R:A B, is a subset of A B. (A 에서 B 로의이진관계
이산수학 () 관계와그특성 (Relations and Its Properties) 2010년봄학기강원대학교컴퓨터과학전공문양세 Binary Relations ( 이진관계 ) Let A, B be any two sets. A binary relation R from A to B, written R:A B, is a subset of A B. (A 에서 B 로의이진관계
sirna 실험은잘됐는데... 그러면이제유전자의기능을완전히억제하고싶은데... 1 화 : CRISPR 은손쉽다 CRISPR 에게맡겨봐! 내게염기서열만알려주면 knock out 을시켜버릴게! Gene editing( 게놈편집 ) 을이용하면특정유전자의기능을완전히 knock-
 을이용하면특정유전자의기능을완전히 knock-") Cover title Subhead GeneArt CRISPR GeneArt Precision TALENs 게놈편집이펼치는과학원더랜드 만화로만나는게놈편집의원리 sirna 실험은잘됐는데... 그러면이제유전자의기능을완전히억제하고싶은데... 1 화 : CRISPR 은손쉽다 CRISPR 에게맡겨봐! 내게염기서열만알려주면 knock out 을시켜버릴게! Gene editing(
Cover title Subhead GeneArt CRISPR GeneArt Precision TALENs 게놈편집이펼치는과학원더랜드 만화로만나는게놈편집의원리 sirna 실험은잘됐는데... 그러면이제유전자의기능을완전히억제하고싶은데... 1 화 : CRISPR 은손쉽다 CRISPR 에게맡겨봐! 내게염기서열만알려주면 knock out 을시켜버릴게! Gene editing(
Jkafm093.hwp
 가정의학회지 2004;25:721-739 비만은 심혈관 질환, 고혈압 및 당뇨병에 각각 위험요인이고 다양한 내과적, 심리적 장애와 연관이 있는 질병이다. 체중감소는 비만한 사람들에 있어 이런 위험을 감소시키고 이들 병발 질환을 호전시킨다고 알려져 있고 일반적으로 많은 사람들에게 건강을 호전시킬 것이라는 믿음이 있어 왔다. 그러나 이런 믿음을 지지하는 연구들은
가정의학회지 2004;25:721-739 비만은 심혈관 질환, 고혈압 및 당뇨병에 각각 위험요인이고 다양한 내과적, 심리적 장애와 연관이 있는 질병이다. 체중감소는 비만한 사람들에 있어 이런 위험을 감소시키고 이들 병발 질환을 호전시킨다고 알려져 있고 일반적으로 많은 사람들에게 건강을 호전시킬 것이라는 믿음이 있어 왔다. 그러나 이런 믿음을 지지하는 연구들은
<4D F736F F F696E74202D203137C0E55FBFACBDC0B9AEC1A6BCD6B7E7BCC72E707074>
 SIMATIC S7 Siemens AG 2004. All rights reserved. Date: 22.03.2006 File: PRO1_17E.1 차례... 2 심벌리스트... 3 Ch3 Ex2: 프로젝트생성...... 4 Ch3 Ex3: S7 프로그램삽입... 5 Ch3 Ex4: 표준라이브러리에서블록복사... 6 Ch4 Ex1: 실제구성을 PG 로업로드하고이름변경......
SIMATIC S7 Siemens AG 2004. All rights reserved. Date: 22.03.2006 File: PRO1_17E.1 차례... 2 심벌리스트... 3 Ch3 Ex2: 프로젝트생성...... 4 Ch3 Ex3: S7 프로그램삽입... 5 Ch3 Ex4: 표준라이브러리에서블록복사... 6 Ch4 Ex1: 실제구성을 PG 로업로드하고이름변경......