특허청구의범위청구항 1 클리오퀴놀 (clioquinol) 또는하기화학식 1의구조를가지는, 그의유도체를유효성분으로포함하는비만예방및억제용약학조성물 : < 화학식 1> R 은 H 또는아세틸기, X 1 또는 X 2 는독립적으로 H 또는할로겐원자. 청구항 2 삭제청구항 3 제
|
|
|
- 동욱 편
- 7 years ago
- Views:
Transcription
 (21) 출원번호 10-2007-0046497 (22) 출원일자 2007 년 05 월 14 일 심사청구일자 2007 년 05 월 14 일 (65) 공개번호 10-2008-0100600 (43) 공개일자 2008 년 11 월 19 일 (56) 선행기술조사문헌 KR1020030027110 A KR1020040036001 A JP02157267 A")
1 (51) Int. Cl. (19) 대한민국특허청 (KR) (12) 등록특허공보 (B1) A61K 31/47 ( ) A61P 3/04 ( ) (21) 출원번호 (22) 출원일자 2007 년 05 월 14 일 심사청구일자 2007 년 05 월 14 일 (65) 공개번호 (43) 공개일자 2008 년 11 월 19 일 (56) 선행기술조사문헌 KR A KR A JP A WO A1 (45) 공고일자 2008년12월22일 (11) 등록번호 (24) 등록일자 2008년12월16일 (73) 특허권자 서울시립대학교산학협력단 서울동대문구전농동 90 (72) 발명자 박현성 경기성남시분당구서현동삼성한신아파트 최수미 서울광진구능동 (74) 대리인 이원희 전체청구항수 : 총 7 항심사관 : 조경주 (54) 클리오퀴놀및그의유도체를유효성분으로함유하는지방형성억제용조성물 (57) 요약 본발명은클리오퀴놀및그의유도체를유효성분으로함유하는지방형성억제용조성물에관한것으로, 지방세포분화시클리오퀴놀의농도의존적이고효과적으로지방형성을억제하며, FAS(fatty acid synthase) 의발현을감소시킴으로지방형성억제및비만치료및예방에유용하게이용될수있다. 대표도 - 1 -
2 특허청구의범위청구항 1 클리오퀴놀 (clioquinol) 또는하기화학식 1의구조를가지는, 그의유도체를유효성분으로포함하는비만예방및억제용약학조성물 : < 화학식 1> R 은 H 또는아세틸기, X 1 또는 X 2 는독립적으로 H 또는할로겐원자. 청구항 2 삭제청구항 3 제 1항에있어서, 할로겐원자는 F, Cl, Br 또는 I인것을특징으로하는비만예방및억제용약학조성물. 청구항 4 제 1항에있어서, 클리오퀴놀의유도체는클로로아세톡시퀴놀린 (chloroacetoxy quinoline : 5- chloroquinolin-8-yl acetate), 브록시퀴놀린 (broxyquinoline : 5,7-diiodo-8-hydroxyquinoline), 5,7-디브로모-8-히드록시퀴놀린 (5,7-dibromo-8-hydroxyquinoline) 또는 8-하이드록시퀴놀 (8-hydroxyquinoline) 인것을특징으로하는비만예방및억제용약학조성물. 청구항 5 제 1항에있어서, 클리오퀴놀또는그의유도체 HIF-1α(hypoxia-inducible factor 1 alpha) 의유비퀴틴화를억제하거나 HIF-1α 전사활성을증대시키는것을특징으로하는비만예방및억제용약학조성물. 청구항 6 클리오퀴놀 (clioquinol) 또는하기화학식 1의구조를가지는그의유도체를유효성분으로포함하는비만예방및억제용건강식품 : < 화학식 1> R 은 H 또는아세틸기, X 1 또는 X 2 는독립적으로 H 또는할로겐원자. 청구항 7-2 -
3 삭제청구항 8 제 6항에있어서, 할로겐원자는 F, Cl, Br 또는 I인것을특징으로하는비만예방및억제용건강식품. 청구항 9 제 6항에있어서, 클리오퀴놀의유도체는클로로아세톡시퀴놀린 (chloroacetoxy quinoline : 5-chloroquinolin- 8-yl acetate), 브록시퀴놀린 (broxyquinoline : 5,7-diiodo-8-hydroxyquinoline), 5,7-디브로모-8-히드록시퀴놀린 (5,7-dibromo-8-hydroxyquinoline) 또는 8-하이드록시퀴놀 (8-hydroxyquinoline) 인것을특징으로하는비만예방및억제용건강식품. 청구항 10 삭제청구항 11 삭제청구항 12 삭제청구항 13 삭제청구항 14 삭제청구항 15 삭제청구항 16 삭제청구항 17 삭제청구항 18 삭제청구항 19 삭제 명세서 발명의상세한설명 발명의목적 <47> 발명이속하는기술및그분야의종래기술 본발명은클리오퀴놀및그의유도체를유효성분으로함유하는지방형성억제용조성물에관한것으로구체적 으로본발명의억제용조성물은지방형성을효과적으로억제하며 FAS(fatty acid synthase) 의발현을 - 3 -
4 저해한다. <48> <49> 저산소상태하에서, 세포는미토콘드리아에의한산화적인산화에필요한산소호흡을유지할수없고그결과로 ATP 생성의감소를가져온다. 산소-관용세포중많은종류는혐기대사 (anaerobic metabolism) 의증가뿐아니라에너지를위한수요감소로인한에너지결핍의위험을해결하였다 (Hochachka, P.W. 1986, Science, 231, ). 많은연구들은상기와같은적응대사 (adaptation mechamism) 가주로해당효소 (glycolytic enzym e) 의활성증가로인한혐기대사에의존함을보여준다. 그와는대조적으로, ATP를위한수요감소또한적응에중요하다는증거들이있다. 산소결핍상태에적응한간세포주에서광범위한단백질합성이신속하게저해되었으며, 이를통해 ATP의수요가감소되었다 (Hochachka, P.W. et al., 1996, Proc. Natl. Acad. Sci. USA 93: ). 이러한결과는저산소세포가 ATP를많이사용하는단백질및지질합성과같은동화과정 (anabolistic process) 을감소시킴을시사하며, 본발명자들은이와같은과정이실제 ATP 레벨의감소이전에일어나서피드포워드 (feedforward) 조절이일어난다고추측하였다. 또한저산소상태에서지방세포분화가억제된다는보고가있다 (Yun. Z. et al., 2002, Dev Cell. 2: ). 저산소상태는 bhlh(basic helix-loop-helix)-pas 전사인자인 HIF-1α(hypoxia-inducible factor 1α) 의레벨증가에의한많은유전자의발현을유도한다. HIF-1α는 HIF-1β(bHLH-PAS 단백질 ) 과기능적이중복합체 (heterodimer) 형태를가진다. HIF-1β는 Arnt(aryl hydrocarbon receptor nuclear tranlocator) 로밝혀졌는데, 이는 AhR(aryl hydrocarbon receptor) 의짝으로밝혀졌다 (Li, H., et al., 1996, J. Biol. Chem, 271: ; Wang, G. L. et al., 1995, J. Biol. Chem. 270: ). 고도로보존된 HIF-1β는안정적인반면, HIF-1α는극심하게불안정하며, 이는산소에의해조절된다. 정상산소상태에서번역된 HIF-1α 단백질은유비퀴틴화되고, 유비퀴틴프로테오좀대사경로에의해신속하게분해된다 (Huang, L. E., et al., 1996, J. Biol. Chem. 271: ; Kallio, P. J., et al., 1999, J., Biol. Chem. 274: ). HIF- 1α는산소-의존적분해도메인을포함하고있으며 (Huang, L. E., et al., 1998, Proc. Natl. Acad. Sci. USA 95: ), 종양억제자인 pvhl(von Hippel-Lindau protein) 과결합하는자리를포함하는고도로보존된지역 (region) 이있다 (Cockman, M. E., et al., 2000, J. Biol. Chem. 275: ). pvhl은분해를위한표적화로 HIF-1α를유비퀴틴화시키는유비퀴틴 E3 리가제 (ligase) 를활성화시키는복합체의조립을구성한다. 최근연구결과는 pvhl이 ODD (HIF-1α의 oxygen dependent degradation domain) 도메인상의보존된프롤린 (proline) 잔기와결함함을제시한다. 산소분자및 Fe 2+ 가필요한과정에서수산화된다 (Ivan, M., et al., 2001, Science 292: ; Jaakkola, P., et al., 2001, Science 292: ). <50> <51> 지방형성 (lipogenesis) 은글루코오스 (glucose) 가몇몇효소에의해트리글리세리드 (triglyceride) 로변하는과정을의미하며, 간및지방조직에서일어난다. 글루코오스는해당과정효소들 (glycolytic enzymes) 에의해아세틸-CoA(acetyl-CoA) 로변환되고, 아세틸-CoA는 ACC(acetyl-CoA carboxylase) 에의해 malonyl-coa로변환되고, malonyl-coa는 FAS(Fatty acid synthase) 에의해지방산 (fatty acid) 으로변하고, 지방산은 glycerol-p-transferase에의해 fatty acyl-coa로변하고, fatty acyl-coa는다른지방형성효소에의해트리글리세리드로변환한다 (Kersten, S., 2001, EMBO, Rep. 2: ). 지방산의내생적합성을책임지는지방형성효소인 FAS(Fatty acid synthase) 는전사및활성레벨에서호르몬및영양촉진에의해조절되는생산물중하나임을확인하였다 (Bennett, M. K., et al., 1995, J. Biol. Chem. 270: ; Wang, Y., et al., 2004, J. Nutr. 134: ). 인슐린및스테롤은 FAS 유전자발현에장기적영향을미치며 (Assimacopoulos-Jeannet, F.S., et al., 1995, Metabolism 44: ), 대개는전사인자, ADD1/SREBP1c(adipocyte determination, and differentiationdependent factor 1/sterol regulatory element binding protein 1c) 를통해서영향을미친다 (Horton, J.D., 1999, J.Clin. Invest. 103: ; Shimomura, I., et al., 1999, J. Biol. Chem. 274: ). ADD1/SREBP1c는 bhlh leucine zipper family에속하며 125 kda 전구체단백질로소포체 (endoplasmic reticulum) 막에결합되어합성된다. ADD1/SREBP1c은스테롤-의존적연속 2단계단백질분해과정에의해절단되어소포체막에서분리되며, 분리된 ADD1/SREBP1c의 N-터미널지역을포함한 68 kda의성숙한전사인자는핵내로이동된다 (Brown, M.S., et al., 1997, Cell, 89: ). 단백질절단에의한핵이동은주로스테롤 (Wang, X., et al., 1994, Cell 77:53-62) 및고도불포화지방산 (polyunsaturated fatty acid) 이감소되면개시된다 (Yahagi, N., et al., 1999, J. Biol. Chem. 274: ). 그밖에도 ADD1/SREBP1c mrna 및단백질의레벨은 AMPK(AMP-activated protein kinase), 금식후재섭취 (Gosmain, Y., et al., 2005, J. Lipid Res. 46: ) 및인슐린 (Chen, G., et al., 2004, Proc. Natl. Acad. Sci. USA, 101: ; Yoshikawa, T., et al., 2001, Mol. Cell. Biol. 21: ) 등의많은신호에의해조절된다. metformin에 - 4 -
5 의한 AMPK 의인산화되어활성화되면 ACC 의인산화시켜활성을억제하고 ADD1/SREBP1c 발현의감소를일으킨다 (Zhou, G., et al., 2001, J. Clin. Invest. 108: ). <52> 클리오퀴놀 (Clioquinol: 이하 "CQ" 라칭함 ) 은 Zn 2+, Cu 2+ 및 Ca 2+ 와같은중금속이온을선택적으로킬레이트화 시키며, 효소활성및단백질형성에있어서중금속이온들의효과를조절하는데사용된다. CQ 의 pka 값은 Cu 2+, 15.8; Zn 2+, 12.5; Ca 2+, 8.1; Mg 2+, 8.6이다 (Agrawal, Y. K. et al., J Pharm Sci. 75: , 1986). CQ는소수성이며, 뇌혈관장벽 (blood brain barrier) 을통과한다. CQ는최근알츠하이머질환, 파킨슨질환, 헌팅턴질환에서메탈로단백질의침전과그로인한산화스트레스를감소시키는 prototype metal-protein attenuating compound로재평가되고있다. CQ는 1900년대중반에항생제로각광받았으나일본에서골수-시각신경병증의원인으로밝혀지면서회수되었다 (Cherny, R. A., Neuron 30: , 2001; Kaur, D. et al., Neuron 37: , 2003; Nguyen T. et al., Proc Natl Acad Sci USA 102: , 2005). 최근알츠하이머질환동물모델인 APP2576 형질전환마우스연구에서 CQ는아밀로이드베타플러그와아밀로이드세럼수치를부작용 (adverse effect) 없이감소시킨다고보고되었다 (Doraiswamy, P. M. et al., Lancet Neorul 3: , 2004). 최근 2단계임상조사에서알츠하이머환자 36명을대상으로한실험에서 CQ는인지감퇴를늦추었으며, 유의적으로아밀로이드베타농도를감소시켰다 (Ritchie C. W. et al., Arch Neurol 60: , 2003). <53> <54> CQ는또한여러인간암세포주에서세포사멸을유발한다. CQ를처리한암세포주에구리 (copper) 또는징크 (zinc) 를첨가하여도 CQ로인한세포죽음은막을수없으며, 오히려더세포죽음을증가시켰다. CQ 처리세포에형광표지로징크수치가증가하는것을관찰하였는데, 이를통해 CQ가이온투과담체 (zinc ionophore) 의활성화에의해세포죽음을일으킴을알수있다 (Ding, W. O., et al., Cancer Res. 65: , 2005). 현재까지 CQ와지방형성과의관계는밝혀진바없다. 이에본발명자들은 HIF-1이지방형성에미치는영향을조사하던중 CQ 및그의유도체가지방형성을저해함을확인함으로써본발명을완성하였다. <55> 발명이이루고자하는기술적과제 본발명의목적은클리오퀴놀또는그의유도체를유효성분으로포함하는지방형성억제용조성물및비만예방 및억제용건강식품을제공하는것이다. <56> <57> <58> <59> <60> <61> 발명의구성및작용상기목적을달성하기위하여, 본발명은클리오퀴놀 (clioquinol) 또는그의유도체를유효성분을포함하는지방형성 (lipogenesis) 억제용조성물을제공한다. 또한, 본발명은클리오퀴놀또는그의유도체를유효성분을포함하는비만예방및억제용건강식품을제공한다. 또한, 본발명은클리오퀴놀또는그의유도체를유효성분으로포함하는 FAS(fatty acid synthase) 억제용조성물을제공한다. 또한, 본발명은 STRA13 단백질을유효성분으로포함하는지방형성억제용조성물을제공한다. 아울러, 본발명은 HIF-1α 단백질을유효성분으로포함하는지방형성억제용조성물을제공한다. 이하, 본발명을상세히설명한다. <62> <63> 본발명자들은저산소상태및정상산소상태하에서지방전구세포인 3T3-L1 세포주를대상으로지방형성을조사하였다. 그결과, 종래공지된바와같이저산소상태일때지방형성이저해됨을확인하였다 ( 도 1 참조 ). 또한저산소상태하에서트리글리세이드및콜레스테롤의형성에미치는영향을조사한결과, 정상산소상태에서보다저산소상태에서트리글리세이드및콜레스테롤의생성이저해됨이관찰되었다 ( 도 2 참조 ). 이후여러세포주에서정상산소상태및저산소상태하에서의 FAS의발현정도를조사하였다. 그결과, 저산소상태에서는 FAS의 mrna 발현이감소함이관찰되었다 ( 도 3a 참조 ). 또한저산소상태하에서의 ATP의양을측정하였다. 그결과, 정상산소상태보다저산소상태하에서 ATP의양이감소함이측정되었다 ( 도 3b의 C 참조 ). 이 - 5 -
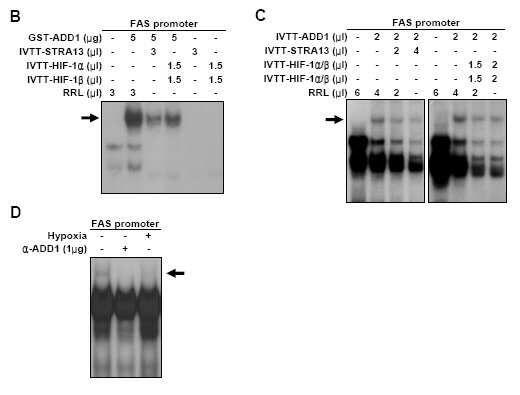
6 를통해궁극적으로저산소상태하에서는 FAS와같은동화효소의감소를통해 ATP 실제감소전에동화작용을막음을알수있었다. 이후본발명자들은 HIF-1β-결핍된세포주를제조하였고, 상기세포주상에서의 FAS 의 mrna 발현량을야생형세포주와비교하였다. 그결과, HIF-1β-결핍된세포주상에서저산소상태로인한 FAS mrna의발현억제현상을상실함을확인하였다. 이는 HIF-1β가 FAS의저산소적발현감소에필수적인분자임을알려주는결과이다. <64> <65> <66> <67> 지방생합성유전자발현은 ADD1/SREBP1c와같은잠재적지방생합성활성인자에의해시작된다. ADD1/SREBP1c는 bhlh-lz(helix-loop-leucine zipper) 패밀리에속한다. real-time PCR을이용하여본발명자들은저산소처리시인간간세포인 HepG2 세포주에서 ADD1/SREBP1c의 mrna양이감소하며 ( 도 4a의 A 참조 ), ADD1/SREBP1c 단백질레벨또한감소함을확인하였다 ( 도 4b의 B 참조 ). 노던분석및 RT-PCR 결과는 ADD1/SREBP1c이야생형세포내에서저산소상태하에서감소됨을나타내나, HIF-1β-결핍 Hepalclc7 세포에서는그렇지않았다. 이를통해 ADD1/SREBP1c의저산소적감소에는 HIF-1β가필요함을알수있다 ( 도 4b의 C 참조 ). 이후, 웨스턴블롯으로 HIF-1β 결핍세포에서는저산소상태가 ADD1/SREBP1c의발현감소에실패함을확인하였다 ( 도 4b의 D 참조 ). 이는 HIF-1β가 ADD1/SREBP1c의저산소적감소에필요함을알려준다 ( 도 4b의 C 및 D 참조 ). FAS의저산소적억제가새로운단백질의합성에대한연관성을조사하기위하여, 본발명자들은저산소노출전에 Hepalclc7 세포및 HIF-1β-결핍 Hepalclc7 세포에 cycloheximide를처리하였다 ( 도 5 참조 ). 저산소세포에서 FAS mrna의레벨은야생형 Hepalclc7 세포에서 cycloheximide 처리하면감소되지않았다. 그러나 HIF-1β 결핍세포주에서는저산소및 cycloheximide 처리군모두에서 FAS의발현이달라지지않았다. 이를통해 FAS의저산소적억제에는새로운단백질의합성이필요함을알수있다. 저산소상태는새로이많은단백질의합성을유도한다. 또한유비퀴틴화를저해함으로서저산소노출 1시간후 HIF-1α 단백질을안정화시킨다 (Miyazaki, K. et al., 2002, J. Biol. Chem. 277: ; Sun, H., and R. Taneja, 2000, Proc. Natl. Acad. Sci. USA 97: ; Yun, Z., et al., Dev. Cell 2: ). 이를통해, HIF-1α/β 이중복합체가전사활성자로서기능하며, 이들의표적유전자의발현을증가킴을알수있다. HIF-1은또한특이적전사억제자의발현을활성화시킨다. 저산소상태에서 basic helix-loop-helix 단백질인 STRA13/DEC1/SHARP2는 HIF-1 의존적으로유도되며, PPARγ2, c-myc, 및 STRA13을포함하는여러유전자발현에억제자로서기능한다 (Azmi, S., et al., 2003, J. Biol. Chem. 278: ; St-Pierre, B., et al., 2022, J. Biol. Chem. 277: ). STRA13은 E-Box에결합하여동형복합체를형성하고표적유전자를억제한다 (St-Pierre, B., et al., 2022, J. Biol. Chem. 277: ). 웨스턴블롯결과는저산소성태가 HIF-1α 및 STRA13 모두의레벨을증가시키며 ( 도 5의 B 및 C 참조 ) STRA13의발현은 HIF-1에의존적임을알수있었다 ( 도 5의 D 참조 ). 실험결과 HIF-1이 ADD1/SREBP1의활성및양을저해함으로, 본발명자들은 HIF-1α 또는 STRA13 존재하의 FAS 프로모터상에서 ADD1/SREBP1c 활성변화를조사하였다. 본발명자들은 ADD1/SREBP1c를발현하는플라스미드와랫트 FAS 유전자의상부부위 (-220 bp ~ +25 bp) 을포함하는 FAS 프로모터리포터플라스미드를함께일시적인형질전환을하였고 ( 도 6a의 A), ADD1/SREBP1c의과발현이리포터유전자의발현을증가시킴을발견하였다 ( 도 6의 B 참조 ). 과발현된 ADD1/SREBP1c의존재시, 저산소상태에서는여전히 FAS 발현을억제하는데이는저산소상태가 ADD1/SREBP1c의양을감소시킬뿐아니라이의활성을억제시킴으로서 FAS의발현을억제함을의미한다. HIF-1α 및 STRA13를 ADD1/SREBP1c를함께형질전환할경우정상세포에서 FAS 발현이감소하는데, 이는 HIF-1α 및 STRA13이 FAS 프로모터상의 ADD1/SREBP1c의활성을저해함을알수있다. ADD1/SREBP1c 프로모터는스테롤반응부위염기서열 (sterol regulatory element complex) 를포함하며, ADD1/SREBP1c은자신의프로모터에결합하여자신의전사를증가시킨다. 본발명자들은 ADD1/SREBP1c를암호화하는플라스미드와마우스 ADD1/SREBP1c 유전자의상부부위 (2.7 kb) 을포함하는리포터플라스미드를함께일시적형질전환을하거나, HIF-1α 또는 STRA13를함께일시적형질전환을한후 ADD1/SREBP1c 프로모터활성의변화를조사하였다 ( 도 6b의 C 참조 ). ADD1/SREBP1c의과발현은리포터유전자의발현을증가시켰다 ( 도 6b의 D 참조 ). ADD1/SREBP1c이과발현하더라도저산소상태에서는 ADD1/SREBP1c 프로모터의활성이감소되었다. HIF- 1α 또는 STRA13과 ADD1/SREBP1c을함께형질전환하면정상상태세포에서보다 ADD1/SREBP1c 프로모터를억제하였는데이는 HIF-1α 및 STRA13이 ADD1/SREBP1c 프로모터를저해함을알수있다 ( 도 6b의 D 참조 ). 본발명자들은 HIF-1α 또는 STRA13을 HepG2 세포에형질전환하여 ADD1 단백질을측정하였다. 도 6b의 E에서볼수있듯이, 정상상태세포에서보다 HIF-1α 또는 STRA13의과발현은 ADD1/SREBP1c의양을저해시킴을보여줌으로써 HIF-1α 및 STRA13이연관되어있음을확인하였다. 저산소상태세포에서 HIF-1α의과발현은 ADD1/SREBP1c를더욱억제시킨다
7 <68> <69> <70> 이후, 본발명자들은 HIF-1α 또는 STRA13이 ADD1/SREBP1c가 DNA에결합하는것을저해시키는지조사하였는데, 이는복합체-의존적으로 E-box 서열 (-CANNTG-) 에결합하는데필요한 bhlh 도메인을가지고있기때문이다 ( 도 7a 의 A 참조 ). ADD1/SREBP1c는두가지종류의 DNA 서열특이성을가지는특이적인 bhlh-leucine zipper 단백질이다. ADD1/SREBP1c은 E-box 서열뿐아니라, SRE(sterol regulatory element) 서열에특이적으로결합한다. 대부분의 SREBPs(sterol regulatory binding protein) 는 SRE를인식하고결합한다. 대조적으로 ADD1/SREBP1c 는 FAS 프로모터및 E-Box를포함하는서열에결합하여이들의프로모터를활성화시킨다 ( 도 6a의 A 및도 6b의 C 참조 )(Magana, M. M., et al., 1997, J. Lipid Res. 38: ; Magna, M. M., and T.F. Osborne. 1996, J. Biol. Chem. 271: ). 2개의독립적이고특이적인 SREBP 결합자리는 FAS 프로모터상의 E-Box 서열양측면에있다 (Magna, M. M., and T.F. Osborne. 1996, J. Biol. Chem. 271: ). 도 7b의 B의시험관내 EMSA 결과에서볼수있듯이, 재조합 GST-ADD1/SREBP1c 융합단백질은 FAS 프로모터의 E-box 포함서열과결합하지만 ( 도 7a의 A 참조 ), 시험관내-번역된 STRA13 동형복합체또는심험관내-번역된 HIF-1α/β 이형복합체는결합하지않았다. 대신에이들의존재는 ADD1/SREBP1c가 FAS 프로모터에결합하는것을막는다 ( 도 7b 의 B 참조 ). 본발명자들은 EMSA를위해시험관내-번역된 ADD1/SREBP1c 단백질을이용하였다. 본발명자들은시험관내-전사된 ADD1/SREBP1c가 FAS 프로모터와결합하나, STRA13 및 HIF-1α 존재시 ADD1/SREBP1c 결합이저해됨을확인하였다 ( 도 7b의 C 참조 ). 본발명자들은또한핵추출물을이용하여, 세포내 FAS 프로모터점유율을조사하였으며, 단백질이 FAS 프로모터와결합함을발견하였다 ( 도 7b의 D 참조 ). ADD1/SREBP1c 항체의첨가는 DNA/ 단백질복합체의소실을일으키는데이는 ADD1/SREBP1c이포함되어있음을나타낸다. 도 7b의 B 및 C의결과와일치하게, 저산소처리는 DNA/ 단백질복합체를포함하는 ADD1/SREBP1c의양을감소시킨다. 이러한발견은저산소-유도된 HIF-1α/β 및 STRA13이 ADD1/SREBP1c가 FAS 프로모터에결합하는것을저해함을나타낸다. ADD1/SREBP1c이자신의프로모터에결합하고전사를활성화한다는사실은저산소상태가 ADD1/SREBP1c가자신의프로모터에결합하는것도방해함으로써 ADD1/SREBP1c의양이저해할수있다는것을나타낸다. ADD1/SREBP1c 프로모터는 SRE 복합체 ( 마우스 ADD1/SREBP1c 유전자의 -92 ~ -56 bp) 를포함하고이들은 ADD1/SREBP1c, 즉자신에의해활성화된다고밝혀졌다 (Amemiya-Kudo, M., et al., 2000, J. Biol. Chem. 275: ; Kim, J.B., et al., 1995, Mol. Cell. Biol. 15: ). SRE 복합체는역시 E-box, SRE(sterol responsive element), NF-Y 및 Spl 자리를가지고있다 ( 도 8a의 A 참조 ). EMSA는 GST-ADD1/SREBP1c가올리고뉴클레오티드를포함하는 SRE 복합체에결합함을보여준다. 흥미롭게도 STRA13 동형복합체및 HIF-1α/β 이형복합체모두 SRE 복합체와결합할수있다 ( 도 8b의 B의아래쪽화살표 ). 실험결과에의하면 STRA13 동형복합체또는 HIF- 1α/β 이형복합체가 ADD1/SREBP1c 프로모터에결합은 E-box 서열또는 SRE 서열에특이적이었다. 본발명자들은표지되지않은변이체올리고뉴클레오티드의초과량을첨가하였다 ( 도 8a의 A 참조 ). 표지되지않은 SRE 변이체올리고뉴클레오티드의첨가는 STRA13 및 HIF-1α/β 결합을파괴하였으며, 표지되지않은 E-box 변이체의첨가는실패하였다. 이러한결과는 STRA13 동형복합체또는 HIF-1α/β 이형복합체는 E-box 서열과결합할수있으나, ADD1/SREBP1c 프로모터상의 SRE 서열과는결합하지않는다는것을증명한다 ( 도 8b의 C 참조 ). 이를통해 STRA13 및 HIF-1α/β는 SRE 복합체내의 E-box와결합하는데 ADD1/SREBP1c와경쟁하는것을알수있다. 저산소상태에의해유도된 HIF-1α 및 STRA13은 ADD1/SREBP1c 단백질과결합하여그로인해이의활성도를저해하는지조사하기위하여본발명자들은 GST pull down assay에서박테리아에서발현된 GST-ADD1/SREBP1c 융합 단백질및 [ 35 P]-표지된 HIF-1α,β, STRA13을이용하였다. 도 9a의 A에서보내주는결과에서는 ADD1/SREBP1c 가 STRA13과강하게결합하였으며, 또한약한수위로 HIF-1α와결합하며, HIF-1β는 ADD1/SREBP1c와의결합하지못하였다. 조-면역침전법 (Co-immunoprecipitation) 으로생체내에서 ADD1/SREBP1c가 STRA13이결합함을확인하였다. 생체내에서 ADD1/SREBP1c는 HIF-1α와매우약하게결합했다 ( 도 9b의 B 참조 ). ADD1/SREBP1c 및 STRA13 또는 HIF-1α가 ADD1/SREBP1c에의하여전사활성을저해하는지확인하기위하여본발명자들은 HepG2 세포에 ADD1/SREBP1c와효모 Gal DNA 결합도메인이융합된단백질을 Gal4 결합자리에의해조절되는프로모터리포터플라스미드와같이형질전환하였다. ADD1/SREBP1c는 Gal4 결합자리에결합하여리포터유전자를증가시켰다. 또한 STRA13 또는 HIF-1α와함께발현할경우리포터유전자의전사를억제시킴을알수있었다. 상기결과는 STRA13 및 HIF-1α가 ADD1과결합하며, ADD1의전사활성화능력을억제함을보여준다 ( 도 9b의 C 참조 ). 한편으로는 HIF-1α는 STRA13을유도하며, STRA13가 ADD1/SREBP1c 프로모터상의 E-box 서열과결합하고 ADD1의발현을억제함을알려준다. 그에더불어 HIF- 1α/β 이형복합체는직접적으로 ADD1/SREBP1c 프라이머상의 E-box와결합하며, E-box에결합하여 ADD1/SREBP1c를저해함을알수있다. <71> 이와같이저산소상태에서지방형성이저해되며, 이에 HIF-1 이관련되어있음을확인한본발명자들은 HIF-1α 의활성을증가시키는클리오퀴놀및그의유도체를이용하여지방형성에미치는결과를조사하였으며그결과 - 7 -
8 는하기에기술하였다. <72> <73> 본발명에서사용하는클리오퀴놀 (clioquinol: 이하 CQ" 라칭함 ) 또는그의유도체는하기의화학식 1 과같은구조를가진다. < 화학식 1> <74> <75> <76> <77> <78> R은 H 또는아세틸기, X 1 또는 X 2 는독립적으로 H 또는할로겐원자. 상기할로겐원자는 F, Cl, Br 또는 I인것이바람직하다. 본발명에서사용하는 CQ( 화학식 2) 의유도체로는 5-chloroquinolin-8-yl acetate( 화학식 3), 5,7-Dibromo-8- hydroxyquinoline( 화학식 4), 8-hydroxyquinoline( 화학식 5) 및 5,7-diiodo-8-hydroxyquinoline( 화학식 6) 등이있으며, 화학식은하기와같으나이에한정되는것은아니며종래에 CQ의유도체로알려진화합물은모두본발명에이용할수있다. < 화학식 2> <79> <80> < 화학식 3> <81> - 8 -
9 <82> < 화학식 4> <83> <84> < 화학식 5> <85> <86> < 화학식 6> <87> <88> <89> <90> <91> 본발명은클리오퀴놀 (clioquinol: 이하 CQ" 라칭함 ) 또는그의유도체를유효성분을포함하는지방형성 (lipogenesis) 억제용조성물을제공한다. 본발명자들은 CQ 및그의유도체가 HIF-1α의활성을촉진함을밝힌바있다 ( 국제출원 PCT/KR2007/000242). 그러나 CQ 및그의유도체와지방형성과의관계는밝혀진바없다. 또한저산소상태에서지방세포분화가억제됨이밝혀졌다 (Yun. Z. et al., 2002, Dev Cell.2: ). 본발명자들은 CQ 및그의유도체가지방형성에미치는영향을조사하였다. 정상산소상태, 저산소상태및 CQ 처리시지방형성을관찰한결과, 종래에보고된바와일치하게정상산소상태에서보다지방형성이현저히저하되었으며, CQ (5 μm) 처리시에도지방형성이저해되었으나저산소상태의세포주에서보다더뛰어난지방형성억제효과를보였다 ( 도 10 참조 ). 또한 CQ를농도별로처리했을때농도의존적으로지방형성이억제됨을확인하였다 ( 도 11 참조 ). 아울러세포내 VEGF의발현을증가시킴과동시에지방형성에관여하는 FAS의발현을감소시므로 ( 도 12 참조 ), 본발명의 CQ 및그의유도체를유효성분으로포함하는조성물은지방형성억제에유용하게이용될수있다. 본발명의 CQ 및그의유도체는그대로또는약학적으로허용가능한염의형태로사용할수있다. 본발명에서사용가능한염으로는약제학적으로허용가능한무독성염이면특별히한정되지않으며, 예를들어, 염산, 황산, 질산, 인산, 불화수소산, 브롬화수소산, 포름산아세트산, 타르타르산, 젖산, 시트르산, 푸마르산, 말레산, 숙신산, 메탈술폰산, 벤젠술폰산, 톨루엔술폰산, 나프탈렌술폰산등의염형태로사용할수있다. 본발명의지방형성억제용조성물은비만예방및치료를위하여단독으로, 또는수술, 호르몬치료, 화학치료및생물학적반응조절제를사용하는방법들과병용하여사용할수있다
10 <92> <93> <94> 본발명의지방형성억제용조성물은상기 CQ 및그의유도체에추가로동일또는유사한기능을나타내는유효성분을 1종이상함유할수있다. 약제학적으로허용가능한담체는식염수, 멸균수, 링거액, 완충식염수, 덱스트로즈용액, 말토덱스트린용액, 글리세롤, 에탄올및이들성분중 1 성분이상을혼합하여사용할수있으며, 필요에따라항산화제, 완충액, 정균제등다른통상의첨가제를첨가할수있다. 본발명의조성물은실제임상투여시에경구및비경구의여러가지제형으로투여될수있는데, 제제화할경우에는보통사용하는충진제, 증량제, 결합제, 습윤제, 붕해제, 계면활성제등의희석제또는부형제를사용하여조제된다. 더나아가당분야의적정한방법으로또는 Remington's Pharmaceutical Science( 최근판 ), Mack Publishing Company, Easton PA에개시되어있는방법을이용하여각질환에따라또는성분에따라바람직하게제제화할수있다. 본발명의조성물은목적하는방법에따라경구투여하거나비경구투여 ( 예를들어, 정맥내, 피하, 복강내또는국소에적용 ) 할수있으며, 투여량은환자의체중, 연령, 성별, 건강상태, 식이, 투여시간, 투여방법, 배설률및질환의중증도등에따라그범위가다양하다. 투약단위는, 예를들면개별투약량의 1, 2, 3 또는 4 배로, 또는 1/2, 1/3 또는 1/4배를함유할수있다. 개별투약량은바람직하기로는유효약물이 1회에투여되는양을함유하며, 이는통상 1일투여량의전부, 1/2, 1/3 또는 1/4배에해당한다. 본발명의지방형성억제용조성물을레트에경구투여하여독성실험을수행한결과, 경구투여독성시험에의한 50% 치사량 (LD 50 ) 은적어도 5 g/kg 이상인안전한물질로판단된다. 유효용량은 mg / kg이고, 바람 직하게는 3 mg / kg이며, 하루 1-3 회투여될수있다. 용량은반드시이에한정되는것은아니고, 환자의상태 및질환의발병정도에따라변할수있다. <95> <96> <97> <98> <99> <100> <101> 또한, 본발명은클리오퀴놀 ( 이하 CQ" 라칭함 ) 또는그의유도체를유효성분을포함하는비만예방및억제용건강식품을제공한다. 본발명의 CQ 및그의유도체를식품첨가물로사용할경우, 상기지방형성억제용조성물을그대로첨가하거나다른식품또는식품성분과함께사용될수있고, 통상적인방법에따라적절하게사용될수있다. 일반적으로, 식품또는음료의제조시에본발명의억제용조성물은원료에대하여 15 중량부이하, 바람직하게는 10 중량부이하의양으로첨가된다. 그러나건강및위생을목적으로하거나또는건강조절을목적으로하는장기간의섭취의경우에는상기양은상기범위이하일수있으며, 안전성면에서아무런문제가없기때문에유효성분은상기범위이상의양으로도사용될수있다. 상기식품의종류에는특별한제한은없다. 본발명의억제용조성물을첨가할수있는식품의예로는육류, 소세지, 빵, 쵸코렛, 캔디류, 스넥류, 과자류, 피자, 라면, 기타면류, 껌류, 아이스크림류를포함한낙농제품, 각종스프, 음료수, 차, 드링크제, 알콜음료및비타민복합제등이있으며, 통상적인의미에서의건강식품을모두포함한다. 본발명의건강음료조성물은통상의음료와같이여러가지향미제또는천연탄수화물등을추가성분으로서함유할수있다. 상술한천연탄수화물은포도당, 과당과같은모노사카라이드, 말토스, 슈크로스와같은디사카라이드, 및덱스트린, 사이클로덱스트린과같은폴리사카라이드, 자일리톨, 소르비톨, 에리트리톨등의당알콜이다. 감미제로서는타우마틴, 스테비아추출물과같은천연감미제나, 사카린, 아스파르탐과같은합성감미제등을사용할수있다. 상기천연탄수화물의비율은본발명의억제용조성물 100 ml당일반적으로약 0.01~0.04 g, 바람직하게는약 0.02~0.03 g 이다. 상기외에본발명의 CQ 및그의유도체는여러가지영양제, 비타민, 전해질, 풍미제, 착색제, 펙트산및그의염, 알긴산및그의염, 유기산, 보호성콜로이드증점제, ph 조절제, 안정화제, 방부제, 글리세린, 알콜, 탄산음료에사용되는탄산화제등을함유할수있다. 그밖에본발명의 CQ 및그의유도체는천연과일주스, 과일주스음료및야채음료의제조를위한과육을함유할수있다. 이러한성분은독립적으로또는조합하여사용할수있다. 이러한첨가제의비율은크게중요하진않지만본발명의억제용조성물 100 중량부당 0.01~0.1 중량부의범위에서선택되는것이일반적이다. 또한, 본발명은클리오퀴놀 ( 이하 CQ" 라칭함 ) 또는그의유도체를유효성분으로포함하는 FAS(fatty acid synthase) 억제용조성물을제공한다. 본발명의 CQ 또는그의조성물은 VEGF의발현을증가시키면서 FAS의발현을감소시킴으로 ( 도 12 참조 ) FAS의발현을억제하는데유용하게이용될수있다
11 <102> <103> <104> <105> <106> <107> 또한, 본발명은 STRA13 단백질을유효성분으로포함하는지방형성억제용조성물을제공한다. 상기 STRA13 단백질은 ADD1/SEREBP1c의활성을감소시키며 ( 도 6), 이를통해 FAS의발현이감소됨으로지방형성억제용조성물및 FAS 억제용조성물로서유용하게이용될수있다. 상기 STRA13 단백질은서열번호 17의아미노산서열을가진다. 아울러, 본발명은 HIF-1α 단백질을유효성분으로포함하는지방형성억제용조성물을제공한다. 상기 HIF-1α 단백질은 ADD1/SEREBP1c의활성을감소시키며 ( 도 6), 이를통해 FAS의발현이감소됨으로지방형성억제용조성물및 FAS 억제용조성물로서유용하게이용될수있다. 상기 HIF-1α 단백질은서열번호 18의아미노산서열을가진다. 이하, 본발명을실시예및제제예에의하여상세히설명한다. 단, 하기실시예및제제예는본발명을구체적으로예시하는것이며, 본발명의내용이실시예및제제예에의해한정되는것은아니다. <108> <109> <110> < 실시예 1> 저산소상태 (hypoxia) 가지방형성 (lipogenesis) 에미치는영향조사본발명자들은지방선구세포 (preadipocyte) 인 3T3-L1 세포주를대상으로저산소상태가지방형성 (lipogenesis) 에미치는영향을조사하였다. 3T3-L1 세포주는 10% FBS가포함된 DMEM(Dulbecco's modified Eagle's midium)(invitrogen, USA) 에서꽉차게배양하였다. 분화를위하여꽉찬상태에서 10% FBS를포함하는 DMEM 배지, 3-isobutyl-1-methylxanthine(500 μm; Sigma, USA), dexamethasone(2 μm; Sigma, USA) 및 insulin(5 μg/ ml ; Sigma, USA) 을 48시간동안처리하였다. 이후정상상태챔버와저산소상태의배양기로옮겨배양하였다. 대조군으로는정상산소상태 (normoxia) 에서배양한세포주를이용하였으며, 실험군으로는저산소상태 (hypoxia) 에서배양한세포주를관찰하였다. 저산소상태노출을위해세포는혐기성배양기 (0.1% O 2 )(Model 1029, Forma Scientific, Inc.) 에서 5% CO 2, 10% O 2, 및 85% N 2 조건으로 37 에서배양하였다. 배양배지는매일교체하였으며, 5 μg / ml인슐린을포함하는 10% FBS를포함하는 DMEM 배지로교체하여주었다 (Seo, J.B. et al., 2004, Mol. Cell. Biol. 24: ). 성숙한지방세포는 PBS로 2회세척하였으며, 10% formaldehyde 로 1시간동안고정하였다. 오일레드 O(isopropanol에 0.5%) 를증류수로희석하였고, 30분간 37 에서고정된세포와함께방치하였다 (Seo, J.B. et al., 2004, Mol. Cell. Biol. 24: ). 이후세포내염색된지질방울은현미경으로확인하였고사진을찍었다. <111> <112> <113> <114> <115> <116> 그결과, 저산소상태에노출된 3T3-L1 세포주에서는정상산소상태에서배양된세포주보다확연히지방세포형성이저해됨이관찰되었다 ( 도 1). < 실시예 2> 저산소상태에서의트리글리세이드 (triglyceride) 및콜레스테롤 (cholesterol) 형성에미치는영향조사 3T3-L1 세포는실시예 1과같이배양및처리하였다. 8일동안배양한후전체세포용해액을준비하여트리글리세리드 (triglyceride) 및콜레스테롤 (cholesterol) 의레벨을 Sigma사 (USA) 에서공급하는정량적효소과정을통하여측정하였다. 트리글리세리드검출을위하여세포용해액을리파제 (lipose) 를포함하는용액 (Sigma) 과반응 (37 에서 10분내지 20분간 ) 시켰으며, 시료의흡광도를분광광도계 (spectrophotometer) 를이용하여 540 nm 에서측정하였다. 콜레스테롤측정을위하여콜레스테롤에스터라아제 (cholesterol esterase) 및콜레스테롤옥시다아제 (cholesterol oxidase) 를포함하는용액 (Sigma) 과반응 (37 에서 10분내지 20분간 ) 시켜 500 nm에서시료의흡광도를측정하였다 (McGowan, M.W., et al., 1983, Clin. Chem. 29: , Rangaswamy, S., et. al., 1997, Circ. Res. 80:37-44). 트리글리세리드및콜레스테롤의양은세포용해액의단백질레벨로평균화하였다. 그결과, 총단백질당트리글리세이드및콜레스테롤의양이정상산소상태에서보다저산소상태에노출된세포주에서억제됨이관찰되었다 ( 도 2). < 실시예 3> 저산소상태에서의 FAS(fatty acid synthase) 의발현양상조사저산소상태하에서 FAS(fatty acid synthase) 의발현양상을조사하기위하여본발명자들은인간간암세포주인 Hep3B 세포주, 마우스골격근육 (skeletal muscle) 세포주인 L6 세포주, 간엽세포주 (mesenchymal cell
12 line) 인 C2C12 세포주및마우스지방선구세포인 3T3-L1 세포주를대상으로 FAS(fatty accid synthase) 및 PGK-1(phospohglycerate kinase-1), GAPDH(glyceraldehyde-3-phosphate dehydrogenase) 및 18S/16S rrna 의발 현을노던블롯방법을이용하여조사하였다. <117> Hep3B 세포주는 10% FBS가포함된 MEM 배지 (Invitrogen, USA) 를이용하여배양하였으며, L6 세포주, C2C12 세포주및 3T3-L1 세포주는 10% FBS가포함된 DMEM 배지를이용하여배양하였으며, 세포들은 100-mm 조직배양플레이트에 70-80% 정도차도록배양하였고, 처리전에 0.5% FBS를포함한각각의배지로 16 내지 24시간동안처리하였다. 이후각각의세포주는저산소조건 (0.1% O 2 ) 에서 16시간동안노출시켰다. 이후전체 RNA를추출하 였다. 전체 RNA는 RNeasy spin column(qiagen, Inc, USA) 을이용하여수득하였다. 전체 RNA는 1% 포름알데히드-아가로오스 (formaldehyde-agarose) 겔을이용하여전기영동을수행하여나일론막으로전이시켰다. 이후 α-[ 32 P]-표지된 cdna와혼성화시켰고세척후에건조하여 Hyperfile MP(Amersham Biosciences, USA) 에감광시켰다 (Hur, E. et al., 2002, Mol. Pharmacol. 62: ). PGK-1(NM_000291) bp, GAPDH (NM_002046) coding region, 18S/28S 는 Ethidium Bromide로염색되어관찰되었다. FAS는 Seo 등이사용한 pcrii vector에들어있는 FAS의 DNA 서열을제한효소 EcoR1으로잘라서정제하여프로브로사용하였다 (Seo J.B, et al., 2004, Mol. Cell. Biol. 24: ). <118> <119> <120> <121> <122> 그결과, 저산소상태를가해주었을때 Hep3B 세포주, L6 세포주, C2C12 세포주및 3T3-L1 세포주상에서 FAS mrna의레벨을저해하였다. 이와는대조적으로 PGK-1 및 GAPDH와같이저산소상태하에서발현이유도되는유전자의 mrna 레벨은증가하였다 ( 도 3a의 A). 야생형마우스간암세포주인 Hepa1c1c7 세포주는 alpha-mem 배지에배양하였다 (Invitrogen, USA). 상기와같은조건으로저산소상태를처리하여 FAS, PGK-1 및 28S/18S rrna의 mrna 발현을조사하였으며, 시간대는 0, 1, 2, 4, 8, 16 및 24 시간대에조사하였다. 그결과, Hepa1c1c7 세포주상에서는저산소처리시 FAS의발현을감소시키며, PGK-1의발현을증가시켰다 ( 도 3a의 B). < 실시예 4> 야생형마우스 Hepa1c1c7 세포주를대상으로한저산소상태에서의 ATP 측정조사본발명자들은 constant-light signal luciferase assay(atp Bioluminescence Assay Kit CLS II) 를통하여정상및저산소상태세포내에서 ATP 생산레벨을검출하는데이용되었다. 야생형마우스 Hepa1c1c7 세포를 35 mm 조직배양플레이트에서 개가되도록 3 개로배양하였으며, 하룻밤동안배양하였다. 16시간이후에, 세포는저산소상태에 6 및 12시간동안노출하였다. 세포를수득하였고, 제조사에서제공하는방법대로분석하였다. 자세하게는 CLS Ⅱ 키트에서제공하는루시퍼레이즈 (luciferase) 용액 25 μl을희석된세포전체수득물에첨가하였고, 루미노미터 (luminometer; Berthold Lumat LB9501) 에서 2s-integration으로측정하였다. 각시료에해당하는 ATP의몰라 (molar) 양은 ATP 기준 (10-4 에서 M ATP) 을기초로하여연관된루시퍼레이즈유닛과대응되게결정하였다. 루시퍼레이즈어세이는 BSA(bovine serum albumin) 를기준으로이용한브래드포드분석을통해결정된전체단백질을농도로평균화하였다. <123> <124> <125> <126> <127> 그결과, 본발명자들은야생형 Hepa1c1c7 세포내에서저산소상태에노출되어 6시간후에 ATP 생산레벨이감소함을확인하였다 ( 도 3b의 A). < 실시예 5> HIF-1β-결핍된 Hepa1c1c7 세포주에서의저산소적저해조사 <5-1> HIF-1β-결핍된 Hepa1c1c7 세포주제조본발명자들은 Whitlock 등이마우스 hepa1c1c7 세포에 benzo(α)pyrene를처리하여돌연변이를유발한후에다이옥신의반응성을상실한돌연변이세포주를분리하는방법대로 HIF-1β-결핍된 Hepa1c1c7 세포주를제작되었다 (Miller, A.G., D.I.Israel, and J.P.Whitlock, Jr. J. Biol. Chem. 258: , 1983). 이중 BpRc1 세포라명명한돌연변이 hepa1c1c7을분석한결과다이옥신수용체결합단백질인 Arnt (HIF-1β와동일 ) 가결여되었음을확인하였다 (Hyunsung P. Ko, et al,, Mol. Cell. Biol. Vol. 10, No. 1, pp , 1996). 본발명자는 BpRc1을 HIF-1β가결여된 hepa1c1c7 세포로사용하였다. 본발명자들은이후 HIF-1β결핍된세포주를웨스턴블롯방법을이용하여정산산소및저산소상태하에서의야생형및 HIF-1β-결핍된세포주상의 HIF-1β의발현을조사하였다. 각각의군에서취한시료를 8% SDS-PAGE 에전기영동을수행하고니트로셀루로오스막 (Schleicher&Schuell Bioscience, USA) 에이동시켰다. 웨스턴블
13 롯수행시 l차항체는 anti-arnt/hif-1β 항체 (BD Parmingen, USA) 를이용하였으며, 2차항체로는마우스-Ig 복합고추냉이퍼옥시데이트 (mouse Ig conjugated horseradish peroxidase)(1:3,000 희석 ) 을이용하였다. 세포주는저산소상태를 6시간동안처리하였다. 그결과, HIF-1β-결핍된세포주는정상산소또는저산소상태하에서 HIF-1β의발현이관찰되지않음을확인하였다 ( 도 3b의 D). <128> <129> <130> <131> <132> <133> <134> <135> <136> <137> <138> <139> <140> <141> <142> <143> <5-2> HIF-1β-결핍된세포주의저산소상태에서의 FAS 발현정도조사본발명자들은 HIF-1β-결핍된세포주를대상으로실시예 3의노던블롯방법대로 FAS의 mrna 발현을조사하였다. 상기세포주에는저산소상태를 16시간동안처리하였다. 그결과, 저산소상태처리시 HIF-1β-결핍된세포주내에서는 FAS의발현감소를관찰할수없었다 ( 도 3b의 E). <5-3> 저산소상태하에서의 ADD1/SREBP1c의발현정도조사본발명자들은인간간암세포주인 hep3b 세포주를대상으로저산소상태하에서의 ADD1/SREBP1c 유전자의발현을 real-time PCR 방법및웨스턴블롯방법으로조사하였다. 상기세포주에저산소상태로 16시간동안처리하고, real-time RT-PCR을수행하기위하여 1 μg전체 RNA를 Lee 등의방법 (Lee, Y.S. et al., 2003, Nucleic Acid Res. 31: ) 으로수득하였다. real-time RT-PCR 수행시 SYBR Green PCR master mix(applied Biosystems) 를이용하여수행하였으며, 각유전자의상대적인활성양은 ABI PRISM 7000(Applied Biosystems) 으로검출하였다. 사용한 PCR 프라이머는하기와같다 : 인간 ADD1/SREBP1c mrna의정방향프라이머 (gcc atg gat tgc act tt: 서열번호 1) 및역방향프라이머 (caa gag agg agc tca atg: 서열번호 2) ; 18S rrna의정방향프라이머 (acc gca gct agg aat aat gga ata: 서열번호 3) 및역방향프라이머 (ctt tcg ctc tgg tcc gtc tt: 서열번호 4). 상기세포주에저산소상태를 6시간동안처리한후, 실시예 5-1과같이웨스턴블롯방법으로수행하였으며, 1 차항체로는 anti-add1/srebp1c 항체 (BD Pharmingen, USA) 를이용하였으며, 2차항체로는마우스-Ig 복합고추냉이퍼옥시데이트 (mouse Ig conjugated horseradish peroxidase)(1:3,000 희석 ) 를이용하였다. 그결과, 저산소처리시 Hep3B 세포주에서는 ADD1/SREBP1c의 mrna의발현이감소하며 ( 도 4a의 A), 그와동시에단백질레벨에서의발현또한감소함을확인하였다 ( 도 4a의 B). < 실시예 6> 저산소상태하에서 FAS 발현억제로인한새로운단백질의합성조사 <6-1> 저산소상태하에서의 FAS 발현억제조사본발명자들은야생형및 HIF-1β-결핍된 Hepa1c1c7 세포주를대상으로 12시간동안저산소상태로처리하였으며, 그중일부는저산소처리전에 30분동안 CHX(cycloheximide) 100 μm로처리하였다. 이후실시예 5-과같은방법으로 RT-PCR를수행하였다. RT-PCR 조건은하기와같다. 95 에서 5분간초기변성과정을거친후, 95 에서 45초동안변성반응, 56 에서 45초동안프라이머결합반응, 72 에서 60초동안길이연장반응을수행하였고, 상기과정을 30회반복하였다. 이때이용한프라이머는하기와같다 : FAS mrna의정방향프라이머 (tgc tcc cag ctg cag gc: 서열번호 5) 및역방향프라이머 (gcc ccg tag ctc tgg gtg ta: 서열번호 6), 18S rrna의정방향프라이머 (acc gca gct agg aat aat gga ata: 서열번호 3) 및역방향프라이머 (ctt tcg ctc tgg tcc gtc tt: 서열번호 4). 그결과, 저산소처리시야생형세포주에서는 CHX 처리에의해자극을받았으나, HIF-1β-결핍된세포주에서는저산소처리및 CHX 처리군모두에서 FAS의발현이변화함을확인하였다 ( 도 5A). <6-2> 저산소상태하에서의 STRA13의발현변화조사본발명자들은 3T3-L1 세포주를대상으로저산소상태를 4시간동안처리하고 STRA13의 mrna 발현레벨을실시예 5-1과같은웨스턴블롯방법으로수행하였다. 1차항체로는항-STRA13 항체 (Abcam, USA) 를이용하였으며, 2차항체로는마우스-Ig 복합고추냉이퍼옥시데이트 (mouse Ig conjugated horseradish peroxidase)(1:3,000 희석 ) 을이용하였다. 또한 3T3-L1 세포주를저산소상태를 16시간동안처리하고실시예 3과같은노던블롯방법으로 STRA13의 mrna 발현레벨을조사하였다. 아울러야생형및 HIF-1β-결핍된 Hepa1c1c7 세포주를 16시간동안저산소상태에서처리한후에, 5-3과같은
14 RT-PCR 방법으로 STRA13의 mrna 발현레벨을조사하였다. 이때이용한프라이머는하기와같다 : 마우스 ADD1/SREBP1c mrna의정방향프라이머 (cac ttc atc aag gca gac tc: 서열번호 7) 및역방향프라이머 (cgg tag cgc ttc tca atg gc: 서열번호 8) <144> <145> <146> <147> 그결과, 저산소상태에서는 HIF-1α 및 STRA13의레벨을모두증가시키며 ( 도 5의 B 및 C), STRA13의발현은 HIF-1에의존적임을알수있다 ( 도 5의 D). < 실시예 7> 저산소상태하에서의 FAS 프로모터와 ADD/SREBP1c 프로모터의조절조사 <7-1> 일시적형질전환루시퍼레이즈어세이 (transient transfection luciferase assay) 본발명자들은인간 (H), 마우스 (M) 및랫트 (R) 의 FAS 프로모터의서열을비교하였다 ( 도 6a의 A). 박스로표지된서열은조절서열로선택된서열들이며, E-box 일치부분 (consensus), hfire, 간장 FAS 인슐린응답요소 (hapatic FAS insulin response element), SREBP, ADD1/SREBP1c 결합자기측면 E-box(binding site flanked E-box), TATA-box를표시하였다. 본발명자들은 pcdna3-hif-1α(human, U22431)(Hunag, L.E., et al., 1998, Proc. Natl. Acad. Sci. USA 95: ), pcdna3.1-stra13(mouse, AF010305), ADD1/SREBP1c의 bhlhleucine zipper domain에 myc-his가표지된 ADD1/SREBP1c cdna(rat, AF286469) 를포함하는 pcdna3- ADD1/SREBP1c (1-403)-myc/hisA(Lee, Y.S., et al., 2003, Nucleic Acids Res. 31: ) 를각각 250 ng 씩 T3-L1 세포주에형질전환하였으며, 동시에 FAS 유전자의상위조절영역 (-220 bp ~ +25 bp) 를포함하는 FAS promoter-driven luciferase reporter 플라스미드 200 ng을함께형질전환하였다. 세포를수득하기전에형질전환된세포주는정상산소상태또는저산소상태 (0.1% O 2 ) 로 16시간동안처리하였다. 형질전환이 후 48시간이지났을때세포수득물을준비하고루시퍼레이즈어세이시스템 (Promega, USA) 을이용하여루미노미터 (luminometer, Turmer TD-20/20, Promega, USA) 로측정하였다. 루시퍼레이즈활성은브레드포드방법에의해측정된전체단백질농도를이용하여평균화하였다. 형질전환효율은 pcho110(β-glactosidase-encoding plasmid) 를함께형질전환하여 β-갈락토시데이즈를측정하여확인하였다 (Hur, E. et al., 2002, Mol. Pharmacol. 62: ). <148> <149> 그결과, ADD1/SERBP1c를과발현시리포터유전자의발현이증가됨이관찰되었다. 또한 ADD1/SREBP1c의존재량이증가할때저산소상태에서는 ADD1/SREBP1c의양을감소시킬뿐만아니라이의활성을억제시킴으로써 FAS의발현을억제하는것으로보인다. 아울러 HIF-1α 및 STRA13를 ADD1/SERBP1c를함께형질전환시킬경우, 정상세포에서는 FAS의발현이감소하는데이는 HIF-1α 및 STRA13이 FAS 프로모터상의 ADD1/SREBP1c의활성을저해하는것임을알수있다 ( 도 6a의 B). ADD1/SREBP1c 프로모터는스테롤조절요소복합체 (sterol regulatory element complex) 를포함하며, ADD1/SREBP1c 자신에의해유도되는지를조사하기위하여, 본발명자들은 ADD1/SREBP1c를암호화하는플라스미드인 pcdna3-add1/srebp1c (1-403)-myc/hisA(Lee, Y.S., et al., 2003, Nucleic Acids Res. 31: ) 250 ng, pcdna3-hif-1α(human, U22431)(Hunag, L.E., et al., 1998, Proc. Natl. Acad. Sci. USA 95: ) 300 ng, pcdna3.1-stra13(mouse, AF010305) 300 ng을각각 T3-L1 세포주에일시적으로형질전환하였으며, 이때마우스 ADD1/SREBP1c 유전자의상류조절영역 (-2.7 kb ~ + 1 bp) 을포함하는 FAS 프로모터리포터플라스미드를함께형질전환하여 HIF-1α 또는 STRA13의존재하에 ADD1/SREBP1c 프로모터상의 ADD1/SREBP1c의활성변화를조사하였다. 세포수득전에정상산소상태또는저산소상태 (0.1% O 2 ) 로 16시간동안처리하였다. 이후상기와같은방법으로루시퍼레이즈어세이를수행하였다. <150> 그결과, ADD1/SREBP1c의과발현은리포터유전자의발현을증가시켰다. 저산소상태에서는과발현된 ADD1/SREBP1c에의한 ADD1/SREBP1c 프로모터의활성화정도를감소시켰다. HIF-1α 또는 STRA13과 ADD1/SREBP1c를함께형질전환하면정상상태세포에서보다 ADD1/SREBP1c에의한프로모터활성화를억제하였는데이를통해 HIF-1α 및 STRA13이 ADD1/SREBP1c에의한프로모터활성화를저해함을알수있다 ( 도 6b의 D). <151> <152> <7-2> 조면역침전 (Co-immunoprecipitation) 조사본발명자들은 293 세포에 pebg-add1/srebp1c를 pcmv-myc-stra13 또는 pcmv-3flag-hif-1α와함께형질전환을수행하였으며, 전체세포추출물은 Hur 등의방법 (Hur. E., et al., 2002, Mol. Pharmacol. 62: ) 으로준비하였다. IP(immunoprecipitation) 는전체세포용해액중 300 μg 시료를 4 에서 30분간항-마우스 IgG(Santa Cruz Biotechnology, Santa Cruz, CA) 1 μg과 20 μl의 0.5% ImmunoPure immobilized protein A/G
15 gel(pierce, Rockford, IL) 과반응하여전처리하였다. 처리된추출물은항-ADD1/SREBP1c 항체 (Dr. Kim, J.B) 2 μg와함께혼합하였다. 이후 0.5 % ImmunoPure immobilized protein A/G gel 15 μl을첨가한후, 혼합물을 4 에서하룻밤동안방치하였다. IP를침전시키고 PBS로 4회세척하여 SDS 시료버퍼로용해하였다. 시료를 5분동안가열하고 7.5% SDS-PAGE 젤에서영동하였고, 단백질은 semi-dry transfer(trans-blot SD; Bio-Rad, Hercules, CA) 를이용하여니트로셀룰로오스막으로이동시켰다. 조-면역침전시킨단백질은항-Myc 항체 (clone 9E10, Boehringer Mannheim) 또는항-FLAG 항체와반응시키고, HRP(horseradish peroxidase) 와결합된항마우스 Ig와함께증강된 chemiluminescence(pierce, Rockford, IL) 로시각화시켰다 (Hur, E., et al., 2002, Mol. Pharmacol. 62: ; Yim, S., et al., 2004, Biochem. Biophys. Res. Commun. 322:9-16). <153> <154> <155> 그결과, 정상상태세포에서보다 HIF-1α 또는 STRA13의과발현된세포에서 ADD1/SREBP1c의양이저해됨을확인함으로써 HIF-1α 및 STRA13이연관되어있음을확인하였다. 저산소상태로처리한세포의경우 HIF-1α의과발현은 ADD1/SREBP1c를더욱억제시켰다 ( 도 6b의 E). < 실시예 8> HIF-1α/β 및 STRA13의존재시 ADD1/SREBP1c의 DNA 결합활성도조사본발명자들은 GST(Glutathione S-transferase) 및 GST-ADD1/SREBP1c(1-403 아미노산 ) 융합단백질을대장균에서발현하였고, 제조사에서제공하는방법으로 glutathione uniflow resin(amersham, USA) 을이용하여정제하였다. pcdna3-add1/srebp1c(1-403)-myc/hisa, pcdna3-ha-hif-1α, pcdna3-hif-1β 또는 pcdna3.1-stra13 1 μg을주형으로이용하여 rabbit reticulocyte lysate(promega, USA) 내에서 IVTT(in vitro transcription and translation) 하여해당단백질을발현하였다. [ 35 S]-표지된 ADD1/SREBP1c, HIF-1α, HIF-1β 또는 STRA13 단백질 (15 μl ) 을 NETN 버퍼 (20 mm Tris (ph8.0), 100 mm NaCl, 1 mm EDTA, 0.5 % Nonidet P-40 및 1 mm PMSF) 500 μl에서 immobilized GST 또는 GST-ADD1/SREBP1c와 4 에서 2시간동안반응시켰다. 단백질은 glutathionuniflow resin과결합하고 4 에서 NETN 버퍼 1 ml로 3회동안세척하였고, SDS 시료버퍼에서가열하여용출하였다. 가열한시료는 SDS-PAGE 및방사선사진촬영 (autoradiography) 을수행하였다 ( 도 7a의 A). <156> 본발명자들은 HepG2 세포를 100-mm 조직배양플레이트에 % 차도록배양하였으며, 0.1 % O 2 에서 6 시간 동안배양하였다. 핵추출물은 Yim 등의방법으로수행하였다 (Yim, S., et al., 2004, Biochem. Biophys. Res. Commun. 322:9-16). rabbit reticulocyte lysate(promega, USA) 에서 pcdna3-add1/srebp1c(1-403)- myc/hisa, pcdna-ha-hif-1α, pcdna-hif-1β 또는 pcdna3.1-stra13 1 μg을주형으로하여 IVTT(in vitro transcription and translation) 을이용하였다. GST(glutathione S-transferase)-ADD1/SREBP1c(1-403 아미노산 ) 융합단백질은대장균 (BL21) 에서발현시켰으며, glytathione uniflow resin을이용하여제조자에서제공하는방법대로정제하였다 (Amersham, USA). FAS 프로모터 (-76 bp~-51 bp) 의서열을포함하는 E-box을위한올리고뉴클레오티드 ( 정방향 : 5 -agt tct gtc agc cca tgt ggc gtg gcc g - 3': 서열번호 9, 역방향 : 5 -gta tcg gcc acg cca cat ggg ctg aca g -3': 서열번호 10)( 도 7a의 A) 및 ADD1/SREBP1c 프로모터야생형서열 (-92 bp ~ -56 bp) 의 SRE 복합체서열을위한올리고뉴클레오티드 ( 정방향 : 5'- tgc tga ttg gcc atg tgc gct cac ccg agg ggc ggg g-3': 서열번호 11, 역방향 : 5'-ccc cgc ccc tcg ggt gag cgc aca tgg cca atc agc a-3': 서열번호 12); SRE 변이체서열 ( 정방향 : 5'-tgc tga ttg gcc atg tgc gct aca ccg agg ggc ggg g -3': 서열번호 13, 역방향 : 5'-ccc cgc ccc tcg gtg tag cgc aca tgg cca atc agc a-3': 서열번호 14); E-box 변이체서열 ( 정방향 : 5'-tgc tga ttg gca aag tgc gct cac ccg agg ggc ggg g-3': 서열번호 15, 역방향 : 5'-ccc cgc ccc tcg ggt gag cgc act ttg cca atc agc a-3': 서열번호 16)(Fig. 8a의 A) 을합성하였다. FAS 프로모터를위한올리고뉴클레오티드및 ADD1/SREBP1c를위한올리고뉴클레오티드야생형프로모터는 α-[ 32 P]-dATP 및크레노우효소로표지하였다 (1.75 pmole). ADD1/SREBP1c-programmed rabbit reticulocyte lysates, 재조합 GST- ADD1/SREBP1c 단백질또는핵추출물은 polydeoxyinosinic-deoxycytidylic acid(1 μg ) 를포함하는 20 μl결합반응버퍼 (10 mm Tris [ph7.5], 50 mm KCl, 2.5 mm EDTA, 0.1% (v/v) Triton X-100, 8.5 % (v/v) glycerol, 1 mm DTT, and 0.1 % (w/v) non-fat milk) 과얼음에서 30분간전반응시켰다 (ref-nar). 표지화된올리고뉴클레오티드 (4x 10 5 cpm, 대략 0.3 pmole) 는 ADD1/SREBP1c-programmed rabbit reticulocyte lysate, recombinant GST- ADD1/SREBP1c protein(5 μg) 또는 nuclear extract(8 μg) 와얼음에서 45 분간반응하였고, 반응혼합물은 4 하에서 6 % PAGE 겔에서분리하였다. EMSA 어세이에서, ADD1/SREBP1c 항체 (1 μg) 는표지화된올리오뉴클레오티드를도입하기전에 4 에서첨가하고 2 시간동안반응하였다 (Kim, J.B. et al., 1998, J. Clin, Invest. 101:1-9). <157> 그결과, 시험관내 - 번역된 STRA13 동형복합체또는시험관내 - 번역된 HIF-1α/β 이형복합체는결합하지않는대
16 신이들의존재는 ADD1/SREBP1cDNA가 DNA에결합하는것을막는다는것을확인하였다 ( 도 7b의 B). 또한, 시험관내-번역된 ADD1/SREBP1c가 FAS 프로모터와결합하나, STRA13 및 HIF-1α 존재시 ADD1/SREBP1c 결합이저해됨을확인하였다 ( 도 7b의 C). 핵추출물을이용하여, 세포내 FAS 프로모터점유율을조사하였으며, 단백질이 FAS 프로모터와결합함을발견하였다. 이결과를통해, ADD1/SREBP1c 항체의첨가는 DNA/ 단백질복합체의소실을일으키는데이는 ADD1/SREBP1c이포함되어있음을알수있었다. 도 7b의 B 및 C의결과와일치하게, 저산소처리는 DNA/ 단백질복합체를포함하는 ADD1/SREBP1c의양을감소시킨다. 이러한발견은저산소-유도된 HIF- 1α/β 및 STRA13이 FAS 프로모터에결합하는 ADD1/SREBP1c를저해함을알수있다. <158> <159> <160> 또한, EMSA 결과, GST-ADD1/SREBP1c가올리고뉴클레오티드를포함하는 SRE 복합체에결합하며, STRA13 동형복합체및 HIF-1α/β 이형복합체모두 SRE 복합체와결합할수있음을확인하였다 ( 도 8b의 B 화살표 ). 실험결과에의하면 ADD1/SREBP1c 프로모터와 STRA13 동형복합체또는 HIF-1α/β 이형복합체사이의결합은 E-box 서열에특이적이었다. 본발명자들은표지되지않은변이체올리고뉴클레오티드의초과량을첨가하였다 ( 도 8a의 A). 표지되지않은 SRE 변이체에올리고뉴클레오티드의첨가는 STRA13 및 HIF-1α/β 결합을파괴하였으며, 표지되지않은 E-box 변이체의첨가는실패하였다. 이러한결과는 STRA13 동형복합체또는 HIF-1α/β 이형복합체는 E-box 서열과결합할수있으나, ADD1/SREBP1c 프로모터상의 SRE 서열과는결합하지않음을나타낸다 ( 도 8b의 C). 이를통해 STAR13 및 HIF-1α/β는 SRE 복합체상의 E-box와결합하는데 ADD1/SREBP1c와경쟁하는것을알수있다. < 실시예 9> bhlh 전사활성사이의결합조사본발명자들은 GST(Glutathione S-transferase) 및 GST-ADD1/SREBP1c(1-403 아미노산 ) 융합단백질을대장균에서발현하였고, 제조사에서제공하는방법으로 glutathione uniflow resin(amersham, USA) 을이용하여정제하였다. pcdna3-add1/srebp1c(1-403)-myc/hisa, pcdna3-ha-hif-1α, pcdna3-hif-1β 또는 pcdna3.1-stra13 1 μg을주형으로이용하여 rabbit reticulocyte lysate (Promega, USA) 내에서 IVTT(in vitro transcription and translation) 하여해당단백질을제조하였다. [ 35 S]-표지된 ADD1/SREBP1c, HIF-1α, HIF-1β 또는 STRA13 단백질 (15 μl ) 을 NETN 버퍼 (20 mm Tris (ph8.0), 100 mm NaCl, 1 mm EDTA, 0.5 % Nonidet P-40 및 1 mm PMSF) 500 μl에서레진에결합된 GST 또는 GST-ADD1/SREBP1c와 4 에서 2시간동안반응시켰다. 단백질은 glutathionuniflow resin과결합하고 4 에서 NETN 버퍼 1 ml로 3회동안세척하였고, SDS 시료버퍼에서가열하여용출하였다. 가열한시료는 SDS-PAGE 및방사선사진촬영 (autoradiography) 을수행하였다. <161> <162> 그결과, ADD1/SREBP1c가 STRA13과강하게결합하였으며, 또한약한수위로 HIF-1α와결합하며, HIF-1β는 ADD1/STRBP1c와의결합이실패하였다 ( 도 9a의 A). 본발명자들은실시예 7-2와같은방법과같이면역침전을수행하였다. 상세하게는 pebg-add1/srebp1c를 pcmv- Myc-STRA13 또는 pcmv-3flag-hif-1α와함께 293 세포주에형질전환하였다. 세포수득전, 6시간동안정상산소상태또는저산소상태 (0.1% O 2 ) 로처리하였다. 이후레진에결합된항-ADD1/SREBP1c 항체 (Dr. Kim. J.B. 제공 ) 를이용하여형질전환된세포용해물을이용하여 IP 를수행하였다. <163> <164> <165> <166> <167> 그결과, 생체내에서 ADD1/SREBP1c가 STRA13과는비교적강하게, HIF-α와매우약하게결합함을확인하였다 ( 도 9b의 B). 이후, 본발명자들은효모의 Gal4 단백질결합염기서열에의해발현되는리포터플라드미드 (gal4-tk-luc) 200 ng을 GAL4-ADD1/SREBB1c 200 ng, pcdna3-stra ng, pcdna3-hif-1α 200 ng와함께 HepG2 세포주에형질전환하였다. 이후실시예 7-1과같은방법으로리포터유전자인루시퍼레이즈의발현을조사하였다. 그결과, Gal4-ADD1/SREBP1c는 Gal4 결합자리에결합하여리포터유전자를증가시킴을확인하였다. 또한 STRA13 또는 HIF-1α와함께발현할경우리포터유전자의전사를억제시킴을알수있었다.( 도 9b의 C). 상기결과를통해 HIF-1α는 STRA13을유도하며, STRA13은 ADD1/SREBP1c 프로모터의 E-box 서열및 ADD1/SREBP1c 단백질상과결합하여 ADD1/SREBP1c이그타겟유전자의전사를활성화하는것을억제시킴을알수있다. 그와더불어 HIF-1α/β 이형복합체역시직접적으로 ADD1/SREBP1c 프로모터상의 E-box와결합하거나약하게는 ADD1/SREBP1c와결합하여 ADD1/SREBP1c의타겟유전자의전사를저해함을알수있었다. < 실시예 10> 지방세포분화에대한클리오퀴놀효과조사지방전구세포 (preadipocyte) 인 3T3-L1 세포주를 10% FBS(fetal bovine serum: Invitrogen, USA) 를포함하는 DMEM 배지 (Invitrogen, USA) 를이용하여플레이트상에가득차도록배양하였다. 이후분화는 10% FBS,
17 3-isobutyl-1-nethylxanthine (500 μm), 덱사메타손 (dexamethasone, 2 μm) 및인슐린 (insulin, 5 μg / ml ) 을포함하는 DMEM 배지에서 8일동안배양하면서수행하였다. 배양배지는 10% FBS 및 5 μg / ml인슐린을포함하는 DMEM 배지로 48시간간격으로교체하였다. 상기분화과정을 8 일간수행하였다. 이때상기세포주를정상산소상태, 저산소상태및클리오퀴놀 (1, 2, 5 μm) 을처리하면서분화를유도하였다. 저산소의경우혐기성배양기 (0.1 % O 2: Model 1029, Forma Scientific, Inc.) 에서 5 % CO 2, 10 % H 2 및 85 % N 2 조건에서 37 에서배양하였다. 클리오퀴놀을처리한군의경우배지를교체할때마다교체배지에클리오퀴놀을재처리하였다. 8 일후성숙한지방세포는 PBS로 2회세척하였고 10% 포름알데하이드 (formaldehyde) 로 1시간동안고정하였다. 오일레드 O(oil red O: 0.5% in isopropanol) 를증류수로희석하여최종 0.3% 로사용한다. 고정된세포플레이트에첨가하여 37 에서 30분간반응시켰다. 세포내염색된지질방울은광학현미경으로시각화하였고사진을찍었다. <168> <169> <170> <171> <172> <173> <174> <175> 그결과, 3T3-L1 세포주에클리오퀴놀 5 μm을처리했을때지질방울의형성이억제됨을확인하였다. 대조군으로서는저산소를처리한 3T3-L1 세포주에서는기존의보고대로지질방울의형성이억제됨이확인되었다 ( 도 10). 본발명자들은클리오퀴놀의여러농도 (1, 2, 5 μm) 를처리했을때클리오퀴놀의농도가증가할수록지방형성의억제가증가함을확인할수있었다 ( 도 11). < 실시예 11> 클리오퀴놀및그의유도체에의한유전자변화조사간세포주인 HepG2 세포주에정상산소상태및저산소상태처리하에서 CQ(50 μm) 및이들의유도체 (5-chloroquinolin-8-yl acetate: 50 μm; 5,7-Dibromo-8-hydroxyquinoline : 5, 10, 50 및 100 μm; 및 8- hydroxyquinoline : 5, 10, 50 및 100 μm) 를 16 시간또는처리시간동안처리하였다. 이후역전사를위해전체 RNA를 RNeasy spin column(qiagen, Inc, USA) 으로분리하였고, 1 μg의전체 RNA를 RT-PCR의주형으로이용하였다. 사용한프라이머로는 FAS(NM_007988) 의증폭을위하여 FAS의정방향프라이머 (5 - tgc tcc cag ctg cag gc - 3 : 서열번호 5) 및역방향프라이머 (5 - gcc ccg tag ctc tgg gtg ta - 3 : 서열번호 6) 를사용하였다. VEGF(AF022375) 의증폭을위하여 VEGF의정방향프라이머 (5 - cca tga act ttc tgc tgt ctt -3 : 서열번호 17) 및역방향프라이머 (5 - atc gca tca ggg gca cac ag -3 : 서열번호 18) 를사용하였다. 대조군으로서 18S rrna(x03205) 의증폭을위하여 18S rrna의정방향프라이머 (5 - acc gca gct agg aat aat gga ata -3 : 서열번호 3) 및역방향프라이머 (5 - ctt tcg ctc tgg tcc gtc tt -3 : 서열번호 4) 를사용하였다. RT-PCR 조건은하기와같다. 95 에서 5분간초기변성과정을거친후, 95 에서 45초동안변성반응, 56 에서 45초동안프라이머결합반응, 72 에서 60초동안길이연장반응을수행하였고, 상기과정을 30회반복하였다. 상기조건으로 30회씩반복하였으며 PCR 생산물을 1% 아가로오스젤에서전기영동하여그결과를확인하였다. 그결과, 클리오퀴놀과그의유도체는 VEGF의발현을증가시키는동시에 FAS의발현을감소시킴을확인하였다 ( 도 12). < 실시예 12> 클리오퀴놀또는그의유도체의경구투여급성독성실험본발명자들은클리오퀴놀또는그의유도체 (5-chloroquinolin-8-yl acetate, 5,7-Dibromo-8-hydroxyquinoline 및 8-hydroxyquinoline) 의급성독성을알아보기위하여하기와같은실험을수행하였다. 6주령의특정병원부재 (SPF) SD계레트를사용하여급성독성실험을실시하였다. 클리오퀴놀또는그의유도체 (5-chloroquinolin-8- yl acetate, 5,7-Dibromo-8-hydroxyquinoline 및 8-hydroxyquinoline) 는각각 0.5% 메틸셀룰로오스용액에현탁하여 5 g/kg/15 ml의용량으로단회경구투여하였다. 실험물질투여후동물의폐사여부, 임상증상, 체중변화를관찰하여혈액학적검사와혈액생화학적검사를실시하였으며, 부검하여육안으로복강장기와흉강장기의이상여부를관찰하였다. 실험결과, 실험물질을투여한모든동물에서특기할만한임상증상이나폐사된동물은없었으며, 체중변화, 혈액검사, 혈액생화학검사, 부검소견등에서도독성변화는관찰되지않았다. 그결과, 레트에서 5 g/kg까지독성변화를나타내지않으며, 경구투여최소치사량 (LD 50 ) 은 5 g/kg 이상인안 전한물질로판명되었다. <176> <177> 하기에본발명의조성물을위한제제예를예시한다. < 제제예 1> : 약학적제제의제조
18 <178> <179> <180> 1. 산제의제조 클리오퀴놀 유당 2 g 1 g <181> <182> 상기의성분을혼합하고기밀포에충진하여산제를제조하였다. 2. 정제의제조 <183> <184> <185> <186> 클리오퀴놀옥수수전분유당스테아린산마그네슘 100 mg 100 mg 100 mg 2 mg <187> <188> 상기의성분을혼합한후, 통상의정제의제조방법에따라서타정하여정제를제조하였다. 3. 캡슐제의제조 <189> <190> <191> <192> 5-chloroquinolin-8-yl acetate 옥수수전분유당스테아린산마그네슘 100 mg 100 mg 100 mg 2 mg <193> <194> 상기의성분을혼합한후, 통상의캡슐제의제조방법에따라서젤라틴캡슐에충전하여캡슐제를제조하였다. 4. 환의제조 <195> <196> <197> <198> 5,7-Dibromo-8-hydroxyquinoline 유당글리세린자일리톨 1 g 1.5 g 1 g 0.5 g <199> <200> 상기의성분을혼합한후, 통상의방법에따라 1 환당 4 g 이되도록제조하였다. 5. 과립의제조 <201> <202> <203> <204> 8-hydroxyquinoline 대두추출물포도당전분 150 mg 50 mg 200 mg 600 mg <205> <206> <207> <208> <209> <210> <211> 상기의성분을혼합한후, 30% 에탄올 100 mg을첨가하여섭씨 60 에서건조하여과립을형성한후포에충진하였다. < 제제예 2> : 식품의제조본발명의클리오퀴놀을포함하는식품들을다음과같이제조하였다. 1. 조리용양념의제조본발명의클리오퀴놀을 20~95 중량부로건강증진용조리용양념을제조하였다. 2. 토마토케찹및소스의제조본발명의클리오퀴놀 0.2~1.0 중량부를토마토케찹또는소스에첨가하여건강증진용토마토케찹또는소스를제조하였다
19 <212> <213> <214> <215> <216> <217> <218> <219> <220> <221> <222> <223> <224> <225> <226> <227> <228> <229> <230> <231> <232> <233> <234> <235> 3. 밀가루식품의제조본발명의클리오퀴놀 0.5~5.0 중량부를밀가루에첨가하고, 이혼합물을이용하여빵, 케이크, 쿠키, 크래커및면류를제조하여건강증진용식품을제조하였다. 4. 스프및육즙 (gravies) 의제조본발명의클리오퀴놀 0.1~5.0 중량부를스프및육즙에첨가하여건강증진용육가공제품, 면류의수프및육즙을제조하였다. 5. 그라운드비프 (ground beef) 의제조본발명의클리오퀴놀 10 중량부를그라운드비프에첨가하여건강증진용그라운드비프를제조하였다. 6. 유제품 (dairy products) 의제조본발명의클리오퀴놀 5~10 중량부를우유에첨가하고, 상기우유를이용하여버터및아이스크림과같은다양한유제품을제조하였다. 7. 선식의제조현미, 보리, 찹쌀, 율무를공지의방법으로알파화시켜건조시킨것을배전한후분쇄기로입도 60 메쉬의분말로제조하였다. 검정콩, 검정깨, 들깨도공지의방법으로쪄서건조시킨것을배전한후분쇄기로입도 60 메쉬의분말로제조하였다. 상기에서제조한곡물류, 종실류및클리오퀴놀을다음의비율로배합하여제조하였다. 곡물류 ( 현미 30 중량부, 율무 15 중량부, 보리 20 중량부 ), 종실류 ( 들깨 7 중량부, 검정콩 8 중량부, 검정깨 7 중량부 ), 클리오퀴놀 (3 중량부 ), 영지 (0.5 중량부 ), 지황 (0.5 중량부 ) < 제제예 3> : 음료의제조 1. 건강음료의제조액상과당 (0.5%), 올리고당 (2%), 설탕 (2%), 식염 (0.5%), 물 (75%) 과같은부재료와클리오퀴놀을균질하게배합하여순간살균을한후이를유리병, 패트병등소포장용기에포장하여건강음료를제조하였다. 3. 야채쥬스의제조본발명의클리오퀴놀 5 g을토마토또는당근쥬스 1,000 ml에가하여건강증진용야채쥬스를제조하였다. 4. 과일쥬스의제조본발명의클리오퀴놀 1 g을사과또는포도쥬스 1,000 ml에가하여건강증진용과일쥬스를제조하였다. <236> 발명의효과본발명의지방형성억제용조성물은지방세포분화시클리오퀴놀의농도의존적으로효과적으로지방형성을억제하며, FAS(fatty acid synthase) 의발현을감소시킴으로지방형성억제및비만치료및예방에유용하게이용될수있다. <1> <2> 도면의간단한설명도 1은저산소상태가지방형성에미치는영향을조사한지방세포사진이다. 도 2는저산소상태가트리글리세이드 (triglyceride) 및콜레스테롤 (cholesterol) 형성에미치는영향을조사한그래프이다
20 <3> <4> <5> <6> <7> <8> <9> <10> <11> <12> <13> <14> <15> <16> <17> <18> <19> <20> <21> <22> <23> <24> <25> <26> <27> <28> <29> <30> <31> <32> <33> 도 3a는저산소상태에서의 FAS(fatty acid synthase) 의발현양상을조사한결과이다 : A : Hep3B, L6, C2C12 및 3T3-L1 상의 FAS 발현정도를노던블롯방법으로분석 ; 및 B : Hepa1c1c7 세포주상에서 FAS 발현정도를노던블롯방법으로분석. 도 3b는야생형 Hepa1c1c7 세포주및 HIF-1β-결핍된 Hepa1c1c7 세포주를대상으로한저산소상태에서의변화조사결과이다 : C : 야생형 Hepa1c1c7 세포주상에서정상산소상태및저산소상태하에서의 ATP 생성정도조사그래프 ; D : HIF-1β-결핍된 Hepa1c1c7 세포주상에서의 HIF-1β 발현정도를웨스턴블롯으로조사한젤사진 ; 및 E : HIF-1β-결핍된 Hepa1c1c7 세포주상에서의 FAS의발현감소조사결과. 도 4a는저산소하에서의 ADD1/SREBP1c 발현정도를조사한결과이다 : A : 정상산소상태및저산소상태하에서의 ADD1/SREBP1c mrna의발현정도를 real-time PCR 방법으로조사한그래프 ; 및 B : 정상산소상태및저산소상태하에서의 ADD1/SREBP1c 단백질의발현정도조사한젤사진. 도 4b는 HIF-1β-결핍된 Hepa1c1c7 세포주를대상으로한 ADD1/SREBP1c mrna의발현정도를조사한결과이다 : C : 노던블롯방법 ; 및 D : 웨스턴블롯방법. 도 5는저산소상태하에서 FAS 발현억제와관련된새로운단백질의합성을조사한결과이다 : A : 야생형및 HIF-1β-결핍된 Hepa1c1c7 세포주상에 Cycloheximide(CHX) 를처리후정상산소상태및저산소상태하에서 FAS mrna의발현을조사한젤사진 ; B : 3T3-L1 세포주상에 STRA13 및 HIF-1α의발현을웨스턴블롯으로조사한젤사진 ; C : 3T3-L1 세포주상에서 STRA13 mrna의발현을조사한젤사진 : 및 D : 야생형및 HIF-1β-결핍된 Hepa1c1c7 세포주상에서 STRA13의 mrna 발현을조사한젤사진. 도 6a는저산소상태하에서의 FAS 프로모터의조절을조사한결과이다 : A : FAS 프로모터-루시퍼레이즈리포터서열 ; 및 B : 정상산소상태및저산소상태하의일시적형질전환루시퍼레이즈어세이그래프. 도 6b는저산소상태하에서의 ADD1/SREBP1c 프로모터의조절을조사한결과이다 : C : ADD1/SREBP1c 프로모터-루시퍼레이즈리포터서열 ; 및 D : 정상산소상태및저산소상태하의일시적형질전환루시퍼레이즈어세이그래프 ; 및 E : 정상산소상태및저산소상태하의웨스턴블롯결과. 도 7a는 HIF-1α, HIF-1β, STRA13 및 ADD1/SREBP1c 의구조와 IVTT 발현양을보여준다. A : HIF-1α, HIF-1β, STRA13 및 ADD1/SREBP1c 의 bhlh 전사인자의도메인구조를표시한다이아그램및 IVTT-ADD1, IVTT-HIF-1α, IVTT-HIF-1β 및 IVTT-ADD1/SREBP1c 제조를확인한젤사진. 도 7b는 HIF-1α/β 및 STRA13 존재하의 ADD1/SREBP1c 와 FAS 프로모터와의결합활성도를조사한결과이다 : B : STRA13, HIF-1α 및 HIF-1β 존재시 GST-ADD1/SREBP1c 와 FAS 프로모터와의결합활성도변화를조사한젤사진 ; C : STRA13, HIF-1α 및 HIF-1β 존재시 IVTT-ADD1/SREBP1c와 FAS 프로모터와의결합활성도를조사한젤사진 ; 및 D : 저산소상태에서세포핵내의 ADD1/SREBP1c 와 FAS 프로모터와의결합활성도를조사한젤사진

21 <34> <35> <36> <37> <38> <39> <40> <41> <42> <43> <44> <45> <46> 도 8a는 ADD1/SREBP1c 프로모터부위의서열이다 : A : ADD1/SREBP1c 프로모터부위의야생형, SRE 변이체, E-box 변이체서열. 도 8b는 HIF-1α/β 및 STRA13 존재하의 ADD1/SREBP1c와 ADD1/SREBP1c 프로모터와의결합활성도를조사한결과이다 : B : ADD1, STRA13, HIF-1α 및 HIF-1β 존재시 ADD1/SREBP1c와ADD1/SREBP1c 프로모터와의결합활성도를조사한젤사진 ; 및 C : ADD1, STRA13, HIF-1α 및 HIF-1β, SRE 변이체및 E-box 변이체존재시 ADD1/SREBP1c와 ADD1/SREBP1c 프로모터와의결합활성도를조사한젤사진. 도 9a는 bhlh 전사활성인자들사이의결합을조사한결과이다 : A : GST pull down system으로조사한젤사진. 도 9b는 bhlh 전사활성인자들사이의결합을조사한결과이다 : B: ADD1, STRA13 및 HIF-1α 존재시발현정도를조면역침강법 (IP) 및웨스턴블롯으로조사한젤사진 ; 및 C: ADD1/SREBP1c, STRA13, HIF-1α 형질전환시리포터유전자의발현을조사한그래프. 도 10은클리오퀴놀 (5 μm) 에의한지방형성억제효과를확인한세포사진이다. 도 11은클리오퀴놀 (1, 2 및 5 μm) 에의한지방형성억제효과를확인한세포사진이다. 도 12는클리오퀴놀및그의유도체에의한 FAS 및 VEGF 발현영향효과를확인한결과이다. 도면 도면

22 도면 2 도면 3a
23 도면 3b 도면 4a

24 도면 4b 도면
25 도면 6a 도면 6b

26 도면 7a 도면 7b 도면 8a


27 도면 8b 도면 9a
28 도면 9b

29 도면

30 도면

31 도면 12 서열목록 <110> UNIVERSITY OF SEOUL FOUNDATION OF INDUSTRY-ACADIMIC COOPERATION <120> Inhibiting composition of lipogenesis containing clioquinol and derivatives thereof <130> 7p <160> 18 <170> KopatentIn 1.71 <210> 1 <211> 17 <212> DNA <213> Artificial Sequence <220> <223> human ADD1/SREBP1c mrna forward primer
32 <400> 1 gccatggatt gcacttt 17 <210> 2 <211> 18 <212> DNA <213> Artificial Sequence <220> <223> human ADD1/SREBP1c mrna reverse primer <400> 2 caagagagga gctcaatg 18 <210> 3 <211> 24 <212> DNA <213> Artificial Sequence <220> <223> 18S rrna forward primer <400> 3 accgcagcta ggaataatgg aata 24 <210> 4 <211> 20 <212> DNA <213> Artificial Sequence <220> <223> 18S rrna reverse primer <400> 4 ctttcgctct ggtccgtctt 20 <210> 5 <211>
33 <212> DNA <213> Artificial Sequence <220> <223> FAS mrna forward primer <400> 5 tgctcccagc tgcaggc 17 <210> 6 <211> 20 <212> DNA <213> Artificial Sequence <220> <223> FAS mrna reverse primer <400> 6 gccccgtagc tctgggtgta 20 <210> 7 <211> 20 <212> DNA <213> Artificial Sequence <220> <223> mouse ADD1/SREBP1c forward primer <400> 7 cacttcatca aggcagactc 20 <210> 8 <211> 20 <212> DNA <213> Artificial Sequence <220> <223> mouse ADD1/SREBP1c reverse primer
34 <400> 8 cggtagcgct tctcaatggc 20 <210> 9 <211> 28 <212> DNA <213> Artificial Sequence <220> <223> E-box oligonucleotide <400> 9 agttctgtca gcccatgtgg cgtggccg 28 <210> 10 <211> 28 <212> DNA <213> Artificial Sequence <220> <223> E-box loigonucleotide <400> 10 gtatcggcca cgccacatgg gctgacag 28 <210> 11 <211> 37 <212> DNA <213> Artificial Sequence <220> <223> SRE complex oligonycleotide <400> 11 tgctgattgg ccatgtgcgc tcacccgagg ggcgggg 37 <210> 12 <211> 37 <212> DNA
35 <213> Artificial Sequence <220> <223> SRE complex oligonucleotide <400> 12 ccccgcccct cgggtgagcg cacatggcca atcagca 37 <210> 13 <211> 37 <212> DNA <213> Artificial Sequence <220> <223> SRE mutant oligonucleotide <400> 13 tgctgattgg ccatgtgcgc tacaccgagg ggcgggg 37 <210> 14 <211> 37 <212> DNA <213> Artificial Sequence <220> <223> SRE mutant oligonucleotide <400> 14 ccccgcccct cggtgtagcg cacatggcca atcagca 37 <210> 15 <211> 37 <212> DNA <213> Artificial Sequence <220> <223> E-box mutant oligonucleotide <400>
36 tgctgattgg caaagtgcgc tcacccgagg ggcgggg 37 <210> 16 <211> 37 <212> DNA <213> Artificial Sequence <220> <223> E-box mutant oligonucleotide <400> 16 ccccgcccct cgggtgagcg cactttgcca atcagca 37 <210> 17 <211> 411 <212> PRT <213> Artificial Sequence <220> <223> STRA13 <400> 17 Met Glu Arg Ile Pro Ser Ala Gln Pro Pro Pro Thr Cys Leu Pro Lys Ala Pro Gly Leu Glu His Gly Asp Leu Ser Gly Met Asp Phe Ala His Met Tyr Gln Val Tyr Lys Ser Arg Arg Gly Ile Lys Arg Ser Glu Asp Ser Lys Glu Thr Tyr Lys Leu Pro His Arg Leu Ile Glu Lys Lys Arg Arg Asp Arg Ile Asn Glu Cys Ile Ala Gln Leu Lys Asp Leu Leu Pro Glu His Leu Lys Leu Thr Thr Leu Gly His Leu Glu Lys Ala Val Val Leu Glu Leu Thr Leu Lys His Val Lys Ala Leu Thr Asn Leu Ile Asp
37 Gln Gln Gln Gln Lys Ile Ile Ala Leu Gln Ser Gly Leu Gln Ala Gly Asp Leu Ser Gly Arg Asn Leu Glu Ala Gly Gln Glu Met Phe Cys Ser Gly Phe Gln Thr Cys Ala Arg Glu Val Leu Gln Tyr Leu Ala Lys His Glu Asn Thr Arg Asp Leu Lys Ser Ser Gln Leu Val Thr His Leu His Arg Val Val Ser Glu Leu Leu Gln Gly Gly Ala Ser Arg Lys Pro Leu Asp Ser Ala Pro Lys Ala Val Asp Leu Lys Glu Lys Pro Ser Phe Leu Ala Lys Gly Ser Glu Gly Pro Gly Lys Asn Cys Val Pro Val Ile Gln Arg Thr Phe Ala Pro Ser Gly Gly Glu Gln Ser Gly Ser Asp Thr Asp Thr Asp Ser Gly Tyr Gly Gly Glu Leu Glu Lys Gly Asp Leu Arg Ser Glu Gln Pro Tyr Phe Lys Ser Asp His Gly Arg Arg Phe Ala Val Gly Glu Arg Val Ser Thr Ile Lys Gln Glu Ser Glu Glu Pro Pro Thr Lys Lys Ser Arg Met Gln Leu Ser Glu Glu Glu Gly His Phe Ala Gly Ser Asp Leu Met Gly Ser Pro Phe Leu Gly Pro His Pro His Gln Pro Pro Phe Cys Leu Pro Phe Tyr Leu Ile Pro Pro Ser Ala Thr Ala Tyr Leu
38 Pro Met Leu Glu Lys Cys Trp Tyr Pro Thr Ser Val Pro Val Leu Tyr Pro Gly Leu Asn Thr Ser Ala Ala Ala Leu Ser Ser Phe Met Asn Pro Asp Lys Ile Pro Thr Pro Leu Leu Leu Pro Gln Arg Leu Pro Ser Pro Leu Ala His Ser Ser Leu Asp Ser Ser Ala Leu Leu Gln Ala Leu Lys Gln Ile Pro Pro Leu Asn Leu Glu Thr Lys Asp <210> 18 <211> 826 <212> PRT <213> Artificial Sequence <220> <223> HIF-1 alpha <400> 18 Met Glu Gly Ala Gly Gly Ala Asn Asp Lys Lys Lys Ile Ser Ser Glu Arg Arg Lys Glu Lys Ser Arg Asp Ala Ala Arg Ser Arg Arg Ser Lys Glu Ser Glu Val Phe Tyr Glu Leu Ala His Gln Leu Pro Leu Pro His Asn Val Ser Ser His Leu Asp Lys Ala Ser Val Met Arg Leu Thr Ile Ser Tyr Leu Arg Val Arg Lys Leu Leu Asp Ala Gly Asp Leu Asp Ile Glu Asp Asp Met Lys Ala Gln Met Asn Cys Phe Tyr Leu Lys Ala Leu Asp Gly Phe Val Met Val Leu Thr Asp Asp Gly Asp Met Ile Tyr Ile
39 Ser Asp Asn Val Asn Lys Tyr Met Gly Leu Thr Gln Phe Glu Leu Thr Gly His Ser Val Phe Asp Phe Thr His Pro Cys Asp His Glu Glu Met Arg Glu Met Leu Thr His Arg Asn Gly Leu Val Lys Lys Gly Lys Glu Gln Asn Thr Gln Arg Ser Phe Phe Leu Arg Met Lys Cys Thr Leu Thr Ser Arg Gly Arg Thr Met Asn Ile Lys Ser Ala Thr Trp Lys Val Leu His Cys Thr Gly His Ile His Val Tyr Asp Thr Asn Ser Asn Gln Pro Gln Cys Gly Tyr Lys Lys Pro Pro Met Thr Cys Leu Val Leu Ile Cys Glu Pro Ile Pro His Pro Ser Asn Ile Glu Ile Pro Leu Asp Ser Lys Thr Phe Leu Ser Arg His Ser Leu Asp Met Lys Phe Ser Tyr Cys Asp Glu Arg Ile Thr Glu Leu Met Gly Tyr Glu Pro Glu Glu Leu Leu Gly Arg Ser Ile Tyr Glu Tyr Tyr His Ala Leu Asp Ser Asp His Leu Thr Lys Thr His His Asp Met Phe Thr Lys Gly Gln Val Thr Thr Gly Gln Tyr Arg Met Leu Ala Lys Arg Gly Gly Tyr Val Trp Val Glu Thr Gln Ala Thr Val Ile Tyr Asn Thr Lys Asn Ser Gln Pro Gln Cys Ile Val
40 Cys Val Asn Tyr Val Val Ser Gly Ile Ile Gln His Asp Leu Ile Phe Ser Leu Gln Gln Thr Glu Cys Val Leu Lys Pro Val Glu Ser Ser Asp Met Lys Met Thr Gln Leu Phe Thr Lys Val Glu Ser Glu Asp Thr Ser Ser Leu Phe Asp Lys Leu Lys Lys Glu Pro Asp Ala Leu Thr Leu Leu Ala Pro Ala Ala Gly Asp Thr Ile Ile Ser Leu Asp Phe Gly Ser Asn Asp Thr Glu Thr Asp Asp Gln Gln Leu Glu Glu Val Pro Leu Tyr Asn Asp Val Met Leu Pro Ser Pro Asn Glu Lys Leu Gln Asn Ile Asn Leu Ala Met Ser Pro Leu Pro Thr Ala Glu Thr Pro Lys Pro Leu Arg Ser Ser Ala Asp Pro Ala Leu Asn Gln Glu Val Ala Leu Lys Leu Glu Pro Asn Pro Glu Ser Leu Glu Leu Ser Phe Thr Met Pro Gln Ile Gln Asp Gln Thr Pro Ser Pro Ser Asp Gly Ser Thr Arg Gln Ser Ser Pro Glu Pro Asn Ser Pro Ser Glu Tyr Cys Phe Tyr Val Asp Ser Asp Met Val Asn Glu Phe Lys Leu Glu Leu Val Glu Lys Leu Phe Ala Glu Asp Thr Glu Ala Lys Asn Pro Phe Ser Thr Gln Asp Thr Asp Leu Asp Leu Glu Met Leu Ala Pro Tyr Ile Pro Met Asp Asp Asp Phe Gln Leu Arg Ser
41 Phe Asp Gln Leu Ser Pro Leu Glu Ser Ser Ser Ala Ser Pro Glu Ser Ala Ser Pro Gln Ser Thr Val Thr Val Phe Gln Gln Thr Gln Ile Gln Glu Pro Thr Ala Asn Ala Thr Thr Thr Thr Ala Thr Thr Asp Glu Leu Lys Thr Val Thr Lys Asp Arg Met Glu Asp Ile Lys Ile Leu Ile Ala Ser Pro Ser Pro Thr His Ile His Lys Glu Thr Thr Ser Ala Thr Ser Ser Pro Tyr Arg Asp Thr Gln Ser Arg Thr Ala Ser Pro Asn Arg Ala Gly Lys Gly Val Ile Glu Gln Thr Glu Lys Ser His Pro Arg Ser Pro Asn Val Leu Ser Val Ala Leu Ser Gln Arg Thr Thr Val Pro Glu Glu Glu Leu Asn Pro Lys Ile Leu Ala Leu Gln Asn Ala Gln Arg Lys Arg Lys Met Glu His Asp Gly Ser Leu Phe Gln Ala Val Gly Ile Gly Thr Leu Leu Gln Gln Pro Asp Asp His Ala Ala Thr Thr Ser Leu Ser Trp Lys Arg Val Lys Gly Cys Lys Ser Ser Glu Gln Asn Gly Met Glu Gln Lys Thr Ile Ile Leu Ile Pro Ser Asp Leu Ala Cys Arg Leu Leu Gly Gln Ser Met Asp Glu Ser Gly Leu Pro Gln Leu Thr Ser Tyr Asp Cys Glu Val Asn Ala Pro Ile Gln Gly Ser Arg Asn Leu Leu Gln Gly Glu
42 Glu Leu Leu Arg Ala Leu Asp Gln Val Asn
ƯÇãû
 '' - 1 - - 2 - - 3 - - 4 - - 5 - ' - 6 - - 7 - - 8 - - 9 - - 10 - - 11 - Super-Bio Co., Ltd. KWON, Suk-Tae Thermostable DNA Polymerase-encoding Gene from Thermus sp. X-1 an d Amino Acid Sequence
'' - 1 - - 2 - - 3 - - 4 - - 5 - ' - 6 - - 7 - - 8 - - 9 - - 10 - - 11 - Super-Bio Co., Ltd. KWON, Suk-Tae Thermostable DNA Polymerase-encoding Gene from Thermus sp. X-1 an d Amino Acid Sequence
α α α α α
 α α α α α α α α 太陰調胃湯加減方 dbdb 마우스 肝에 대한 아디포사이토카인 및 발현에 미치는 영향 SREBPs 와 섞어서 하고 마커 또한 하여 전기 영동을 하였다 전기영동을 한 후에 에 를 쪼여서 각 를 확인하였다 이 를 프로그램을 이용해 수치화하 여 분석하였다 肝 조직 동결 절편 분리한 조직은 로 시간 동안 고정 시킨 후 에 세척한 후 물기를
α α α α α α α α 太陰調胃湯加減方 dbdb 마우스 肝에 대한 아디포사이토카인 및 발현에 미치는 영향 SREBPs 와 섞어서 하고 마커 또한 하여 전기 영동을 하였다 전기영동을 한 후에 에 를 쪼여서 각 를 확인하였다 이 를 프로그램을 이용해 수치화하 여 분석하였다 肝 조직 동결 절편 분리한 조직은 로 시간 동안 고정 시킨 후 에 세척한 후 물기를
- 2 -
 - 2 - - 3 - - 4 - ➀ - 5 - - 6 - α - 7 - - 8 - - 9 - - 10 - - 11 - - 12 - - 13 - - 14 - - 15 - - 16 - - 17 - - 18 - - 19 - - 20 - μ - 21 - ➀ - 22 - - 23 - - 24 - α - 25 - - 26 - - 27 - μ - 28 - μ μ - 29
- 2 - - 3 - - 4 - ➀ - 5 - - 6 - α - 7 - - 8 - - 9 - - 10 - - 11 - - 12 - - 13 - - 14 - - 15 - - 16 - - 17 - - 18 - - 19 - - 20 - μ - 21 - ➀ - 22 - - 23 - - 24 - α - 25 - - 26 - - 27 - μ - 28 - μ μ - 29
F. Sequencing FSequencing 01. Sequencing Service DNA sequencing Phone: (ext.4 4)
") FSequencing 01. DNA sequencing Phone: 1588-9788 (ext.4 4) E-mail: [email protected] 01. 242 FAQs 247 개요 상세 설명 바이오니아는 국내 최초로 를 시작하였으며, 매년 개선된 high-throughput sequencing service를 제공하고 있습니다. Plasmid
FSequencing 01. DNA sequencing Phone: 1588-9788 (ext.4 4) E-mail: [email protected] 01. 242 FAQs 247 개요 상세 설명 바이오니아는 국내 최초로 를 시작하였으며, 매년 개선된 high-throughput sequencing service를 제공하고 있습니다. Plasmid
2016 학년도약학대학면접문제해설 문제 2 아래의질문에 3-4분이내로답하시오. 표피성장인자수용체 (epidermal growth factor receptor, EGFR) 는수용체티로신인산화효소군 (receptor tyrosine kinases, RTKs) 의일종으로서세
 는수용체티로신인산화효소군 (receptor tyrosine kinases, RTKs) 의일종으로서세") 본문제에대한지적소유권은동국대학교에있습니다. 본교의서면허락없이무단으로출판, 게재, 사용할수없습니다. 문제 2 2016 학년도약학대학면접문제 아래의질문에 3-4 분이내로답하시오. 표피성장인자수용체 (epidermal growth factor receptor, EGFR) 는수용체티로신 인산화효소군 (receptor tyrosine kinases, RTKs) 의일종으로서세포의생존과증식
본문제에대한지적소유권은동국대학교에있습니다. 본교의서면허락없이무단으로출판, 게재, 사용할수없습니다. 문제 2 2016 학년도약학대학면접문제 아래의질문에 3-4 분이내로답하시오. 표피성장인자수용체 (epidermal growth factor receptor, EGFR) 는수용체티로신 인산화효소군 (receptor tyrosine kinases, RTKs) 의일종으로서세포의생존과증식
이 발명을 지원한 국가연구개발사업 과제고유번호 KGM2420913 부처명 교육과학기술부 연구관리전문기관 연구사업명 전북분원운영사업 연구과제명 저탄소 녹생성장을 위한 바이오매스/에너지 개발 주관기관 한국생명공학연구원 연구기간 2009년 01월 01일 ~ 2009년 12월
 (19) 대한민국특허청(KR) (12) 등록특허공보(B1) (45) 공고일자 2015년04월10일 (11) 등록번호 10-1509796 (24) 등록일자 2015년04월01일 (51) 국제특허분류(Int. Cl.) A61K 36/45 (2006.01) A23L 1/30 (2006.01) A61P 3/04 (2006.01) (21) 출원번호 10-2009-0108173
(19) 대한민국특허청(KR) (12) 등록특허공보(B1) (45) 공고일자 2015년04월10일 (11) 등록번호 10-1509796 (24) 등록일자 2015년04월01일 (51) 국제특허분류(Int. Cl.) A61K 36/45 (2006.01) A23L 1/30 (2006.01) A61P 3/04 (2006.01) (21) 출원번호 10-2009-0108173
특허청구의범위청구항 1 나한과 ((Siraitia grosvenorii) 를 2중건조후절단하는제 1단계 : 상기제 1단계를거쳐얻어진건조물을추출용매로가열추출하는제 2단계 : 상기추출된침출액을 1차여과하는제 3단계 : 상기 1차여과된침출액을 1차농축하는제 4단계 : 상기농
 를 2중건조후절단하는제 1단계 : 상기제 1단계를거쳐얻어진건조물을추출용매로가열추출하는제 2단계 : 상기추출된침출액을 1차여과하는제 3단계 : 상기 1차여과된침출액을 1차농축하는제 4단계 : 상기농") (19) 대한민국특허청 (KR) (12) 공개특허공보 (A) (51) 국제특허분류 (Int. Cl.) A61K 36/42 (2006.01) A61K 9/16 (2006.01) A61K 9/20 (2006.01) (21) 출원번호 10-2013-0052506 (22) 출원일자 2013 년 05 월 09 일 심사청구일자 없음 (11) 공개번호 10-2014-0133016
(19) 대한민국특허청 (KR) (12) 공개특허공보 (A) (51) 국제특허분류 (Int. Cl.) A61K 36/42 (2006.01) A61K 9/16 (2006.01) A61K 9/20 (2006.01) (21) 출원번호 10-2013-0052506 (22) 출원일자 2013 년 05 월 09 일 심사청구일자 없음 (11) 공개번호 10-2014-0133016
목차 1. 서론 줄기세포의간세포분화능평가시고려사항 간세포분화능평가시험법 분
 세포치료제시험정보집 7 줄기세포치료제의 간세포분화능평가시험 2015. 4. 30. 식품의약품안전평가원 차세대줄기세포기반제제평가연구사업단 목차 1. 서론 --------------------------------------------- 1 2. 줄기세포의간세포분화능평가시고려사항 --------------- 2 3. 간세포분화능평가시험법 -----------------------------
세포치료제시험정보집 7 줄기세포치료제의 간세포분화능평가시험 2015. 4. 30. 식품의약품안전평가원 차세대줄기세포기반제제평가연구사업단 목차 1. 서론 --------------------------------------------- 1 2. 줄기세포의간세포분화능평가시고려사항 --------------- 2 3. 간세포분화능평가시험법 -----------------------------
Chapter 26
 11 주 RNA 합성 11.1 DNA-dependent synthesis of RNA: Bacteria에서의 transcription l RNA polymerase (5 subunits로구성 : 2α, β, β, σ) l 효소에의한 RNA 합성의특징 - Complementary sequence to template DNA - RNA chain의합성방향 : 5
11 주 RNA 합성 11.1 DNA-dependent synthesis of RNA: Bacteria에서의 transcription l RNA polymerase (5 subunits로구성 : 2α, β, β, σ) l 효소에의한 RNA 합성의특징 - Complementary sequence to template DNA - RNA chain의합성방향 : 5
½Ç°ú¸Ó¸®¸»¸ñÂ÷ÆDZÇ(1-5)¿Ï
¿Ï") 실과056-094 2013.1.9 7:22 PM 페이지67 MDPREP_RipControl 2007 개정 5학년 검정 지도서 각론 알짜 정리 67 영양소 힘을 내는 일(탄수화물/지방/단백질) 몸의 조직 구성(지방/단백질/무기질/물) 몸의 기능 조절(단백질/무기질/비타민/물) 식품 구성 자전거의 식품과 영양소 식품군 곡류 탄수화물 우리가 활동하는데 필요한 힘을
실과056-094 2013.1.9 7:22 PM 페이지67 MDPREP_RipControl 2007 개정 5학년 검정 지도서 각론 알짜 정리 67 영양소 힘을 내는 일(탄수화물/지방/단백질) 몸의 조직 구성(지방/단백질/무기질/물) 몸의 기능 조절(단백질/무기질/비타민/물) 식품 구성 자전거의 식품과 영양소 식품군 곡류 탄수화물 우리가 활동하는데 필요한 힘을
45(3)-10(048)p fm
-10(048)p fm") The Korean Journal of Microbiology, Vol. 45, No. 3, September 009, p. 86-90 Copyright 009, The Microbiological Society of Korea s Semi-quantitative RT-PCR w v H9N y» ù» 1 Á Á 3 Á 4 Á x 5 Á 5 Á 5 Á 5 Á«x
The Korean Journal of Microbiology, Vol. 45, No. 3, September 009, p. 86-90 Copyright 009, The Microbiological Society of Korea s Semi-quantitative RT-PCR w v H9N y» ù» 1 Á Á 3 Á 4 Á x 5 Á 5 Á 5 Á 5 Á«x
DBPIA-NURIMEDIA
 Original Article Journal of Apiculture 31(2) : 121~131 (2016) The Most Rapid Detection Method against Korean Sacbrood Virus using Ultra-Rapid Reverse-Transcription Real-Time PCR (URRTRT-PCR) Sang-Hyun
Original Article Journal of Apiculture 31(2) : 121~131 (2016) The Most Rapid Detection Method against Korean Sacbrood Virus using Ultra-Rapid Reverse-Transcription Real-Time PCR (URRTRT-PCR) Sang-Hyun
Journal of Life Science 2011, Vol. 21. No μ μ
 Journal of Life Science 2011 Vol. 21. No. 8. 1120~1126 ISSN : 1225-9918 DOI : http://dx.doi.org/10.5352/jls.2011.21.8.1120 μ μ μ α β Journal of Life Science 2011, Vol. 21. No. 8 1121 μ μ 1122 생명과학회지 2011,
Journal of Life Science 2011 Vol. 21. No. 8. 1120~1126 ISSN : 1225-9918 DOI : http://dx.doi.org/10.5352/jls.2011.21.8.1120 μ μ μ α β Journal of Life Science 2011, Vol. 21. No. 8 1121 μ μ 1122 생명과학회지 2011,
Lehninger Principles of Biochemistry 5/e
 CHAPTER 17 Fatty Acid Catabolism 지방산의이화 1. digestion, mobilization, transport of fats ( 지방의소화, 방출, 운반 ) 2. Oxidation of fatty acids ( 지방산산화 ) 3. ketone bodies ( 케톤체 ) 2008 W. H. Freeman and Company Processing
CHAPTER 17 Fatty Acid Catabolism 지방산의이화 1. digestion, mobilization, transport of fats ( 지방의소화, 방출, 운반 ) 2. Oxidation of fatty acids ( 지방산산화 ) 3. ketone bodies ( 케톤체 ) 2008 W. H. Freeman and Company Processing
최민지 / 세계무역기구과 / :16:24-2 -
 - 2 - - 3 - - 4 - - 5 - - 6 - - 7 - - 8 - - 9 - - 10 - - 11 - - 12 - - 13 - - 14 - - 15 - - 16 - - 17 - - 18 - 안식향산안식향산나트륨안식향산칼륨안식향산칼슘 파라옥시안식향산메틸파라옥시안식향산에틸 0.6 이하 ( 안식향산으로서, 파라옥시안식향산에틸또는파라옥시안식향산메틸과병용할때에는안식향산으로서사용량과파라옥시안식향산으로서사용량의합계가
- 2 - - 3 - - 4 - - 5 - - 6 - - 7 - - 8 - - 9 - - 10 - - 11 - - 12 - - 13 - - 14 - - 15 - - 16 - - 17 - - 18 - 안식향산안식향산나트륨안식향산칼륨안식향산칼슘 파라옥시안식향산메틸파라옥시안식향산에틸 0.6 이하 ( 안식향산으로서, 파라옥시안식향산에틸또는파라옥시안식향산메틸과병용할때에는안식향산으로서사용량과파라옥시안식향산으로서사용량의합계가
추가로 본 발명은 상기 형질전환 복제 소로부터 유래된 정육(meat) 및 가공식품에 관한 것이다. 대표도 도 2 색인어 광우병, 프리온, 핵 이식란, 형질전환 복제 소 명세서 도면의 간단한 설명 도 1은 마우스의 PrP 유전자 및 단백질의 모식도를 나타낸 것이다. 도
 및 가공식품에 관한 것이다. 대표도 도 2 색인어 광우병, 프리온, 핵 이식란, 형질전환 복제 소 명세서 도면의 간단한 설명 도 1은 마우스의 PrP 유전자 및 단백질의 모식도를 나타낸 것이다. 도") (51) Int. Cl. 7 C12N 5/16 (19)대한민국특허청(KR) (12) 공개특허공보(A) (11) 공개번호 (43) 공개일자 10-2005-0055937 2005년06월14일 (21) 출원번호 10-2003-0089000 (22) 출원일자 2003년12월09일 (71) 출원인 재단법인서울대학교산학협력재단 서울특별시 관악구 봉천동 산 4-2 (72)
(51) Int. Cl. 7 C12N 5/16 (19)대한민국특허청(KR) (12) 공개특허공보(A) (11) 공개번호 (43) 공개일자 10-2005-0055937 2005년06월14일 (21) 출원번호 10-2003-0089000 (22) 출원일자 2003년12월09일 (71) 출원인 재단법인서울대학교산학협력재단 서울특별시 관악구 봉천동 산 4-2 (72)
농림수산식품부장관귀하 이보고서를 팥의대사성질환개선및기능성규명 에관한연구의최종보고서로제출 합니다 년 2 월 11 일 - 1 -
 농림수산식품부장관귀하 이보고서를 팥의대사성질환개선및기능성규명 에관한연구의최종보고서로제출 합니다. 2013 년 2 월 11 일 - 1 - 연구기관명 : 부산대학교 연구책임자 : 정명호 연구원 : 손용해 연구원 : 장민경 - 2 - Ⅰ. 제목 팥의대사성질환개선및기능성규명 Ⅱ. 연구개발의목적및필요성 팥의대사성질환개선소재발굴과그기능성을과학적으로증명하기위해, 팥의당뇨,
농림수산식품부장관귀하 이보고서를 팥의대사성질환개선및기능성규명 에관한연구의최종보고서로제출 합니다. 2013 년 2 월 11 일 - 1 - 연구기관명 : 부산대학교 연구책임자 : 정명호 연구원 : 손용해 연구원 : 장민경 - 2 - Ⅰ. 제목 팥의대사성질환개선및기능성규명 Ⅱ. 연구개발의목적및필요성 팥의대사성질환개선소재발굴과그기능성을과학적으로증명하기위해, 팥의당뇨,
(72) 발명자 정종수 서울특별시 서대문구 모래내로 319, 101동 405호 (홍은동, 진흥아파트) 김정환 서울특별시 구로구 구로동로21길 7 (구로동) - 2 -
 발명자 정종수 서울특별시 서대문구 모래내로 319, 101동 405호 (홍은동, 진흥아파트) 김정환 서울특별시 구로구 구로동로21길 7 (구로동) - 2 -") (19) 대한민국특허청(KR) (12) 등록특허공보(B1) (51) 국제특허분류(Int. Cl.) B01J 23/34 (2006.01) B01J 37/02 (2006.01) B01J 37/08 (2006.01) B01D 53/86 (2006.01) (21) 출원번호 10-2010-0098306 (22) 출원일자 2010년10월08일 심사청구일자 2010년10월08일
(19) 대한민국특허청(KR) (12) 등록특허공보(B1) (51) 국제특허분류(Int. Cl.) B01J 23/34 (2006.01) B01J 37/02 (2006.01) B01J 37/08 (2006.01) B01D 53/86 (2006.01) (21) 출원번호 10-2010-0098306 (22) 출원일자 2010년10월08일 심사청구일자 2010년10월08일
Glucose pyruvate (EMP경로, HMP 경로, ED 경로 ) 1 Embden-Meyerhof-Parnas pathways(emp 경로 ) - 전체적인반응 : Glucose 2 pyruvate + 2ATP + 2NADH 지방과인지질합성
 1 Embden-Meyerhof-Parnas pathways(emp 경로 ) - 전체적인반응 : Glucose 2 pyruvate + 2ATP + 2NADH 지방과인지질합성") Chapter 7. 미생물대사 (Microbial Metabolism) 탄수화물대사 : EMP경로, HMP경로, ED경로, TCA cycle, Glyoxylate 회로 지방대사 : β-산화, 지방산합성 ( 중성지방, phospholipid 단백질대사 : transamination, decarboxylation 전자전달과산화적인산화반응 * ATP 생성경로 :
Chapter 7. 미생물대사 (Microbial Metabolism) 탄수화물대사 : EMP경로, HMP경로, ED경로, TCA cycle, Glyoxylate 회로 지방대사 : β-산화, 지방산합성 ( 중성지방, phospholipid 단백질대사 : transamination, decarboxylation 전자전달과산화적인산화반응 * ATP 생성경로 :
- 2 -
 - 1 - - 2 - - 3 - - 4 - μ μ μ μ μ μ μ α μ μ μ μ μ - 5 - μ μ μ μ - 6 - μ - 7 - - 8 - - 9 - - 10 - - 11 - - 12 - - 13 - μ μ μ α μ λ - 14 - μ μ μ μ μ - 15 - - 16 - - 17 - - 18 - - 19 - - 20 - - 21 - - 22
- 1 - - 2 - - 3 - - 4 - μ μ μ μ μ μ μ α μ μ μ μ μ - 5 - μ μ μ μ - 6 - μ - 7 - - 8 - - 9 - - 10 - - 11 - - 12 - - 13 - μ μ μ α μ λ - 14 - μ μ μ μ μ - 15 - - 16 - - 17 - - 18 - - 19 - - 20 - - 21 - - 22
<BBFDC8ADC7D02E687770>
 대상본과 1 학년담당교수손정원 2014. 4. 1. 3/4 교시장소 1 의학관제 1 강의실 2. 수업주제 단백질 folding, targeting, 분해 1) 단백질 folding의기본동력에대해서이해한다. 2) chaperone의종류와작용기전에대하여설명한다. 3) proteasome의구조와 proteasome에의한단백질분해에대하여설명한다. 4) ER targeting
대상본과 1 학년담당교수손정원 2014. 4. 1. 3/4 교시장소 1 의학관제 1 강의실 2. 수업주제 단백질 folding, targeting, 분해 1) 단백질 folding의기본동력에대해서이해한다. 2) chaperone의종류와작용기전에대하여설명한다. 3) proteasome의구조와 proteasome에의한단백질분해에대하여설명한다. 4) ER targeting
 1607 - 1 - - 2 - - 3 - 그림 1-4 - - 5 - 그림 3 농도에따른댓잎추출액의항세균효과 - 6 - 그림 4 한천확산법 그림 5 배지에균뿌리는과정 그림 6 배지에균스프레딩하는과정 표 1 실험에사용한미생물의종류와배양에사용한배지 Bacterial Strain Gram (+) bacteria Rothia dentocariosa G1201 Streptococcus
1607 - 1 - - 2 - - 3 - 그림 1-4 - - 5 - 그림 3 농도에따른댓잎추출액의항세균효과 - 6 - 그림 4 한천확산법 그림 5 배지에균뿌리는과정 그림 6 배지에균스프레딩하는과정 표 1 실험에사용한미생물의종류와배양에사용한배지 Bacterial Strain Gram (+) bacteria Rothia dentocariosa G1201 Streptococcus
<30322EBABBB9AE2E687770>
 대한내과학회지 : 제 79 권부록 2 호 2010 청람연구비결과보고 대장암발생과정에서 Prostaglandin 의조절기전규명및암예방약제의반응성표지자로 15-hydroxyprostaglandin dehydrogenase의역할 울산대학교의과대학내과학교실 명승재 서 론 2. 병리조직학적분석 (Histopathologic analysis) 과제는인체에서대장암병기의각단계에서
대한내과학회지 : 제 79 권부록 2 호 2010 청람연구비결과보고 대장암발생과정에서 Prostaglandin 의조절기전규명및암예방약제의반응성표지자로 15-hydroxyprostaglandin dehydrogenase의역할 울산대학교의과대학내과학교실 명승재 서 론 2. 병리조직학적분석 (Histopathologic analysis) 과제는인체에서대장암병기의각단계에서
Jkbcs016(92-97).hwp
.hwp") Expression of bcl-2 and Apoptosis and Its Relationship to Clinicopathological Prognostic Factors in Breast Cancer - A Study with Long Term Follow-up correlated with the survival rate.(journal of Korean
Expression of bcl-2 and Apoptosis and Its Relationship to Clinicopathological Prognostic Factors in Breast Cancer - A Study with Long Term Follow-up correlated with the survival rate.(journal of Korean
특허청구의 범위 청구항 1 복수개의 프리캐스트 콘크리트 부재(1)를 서로 결합하여 연속화시키는 구조로서, 삽입공이 형성되어 있고 상기 삽입공 내면에는 나사부가 형성되어 있는 너트형 고정부재(10)가, 상기 프리캐스 트 콘크리트 부재(1) 내에 내장되도록 배치되는 내부
를 서로 결합하여 연속화시키는 구조로서, 삽입공이 형성되어 있고 상기 삽입공 내면에는 나사부가 형성되어 있는 너트형 고정부재(10)가, 상기 프리캐스 트 콘크리트 부재(1) 내에 내장되도록 배치되는 내부") (19) 대한민국특허청(KR) (12) 등록특허공보(B1) (51) 국제특허분류(Int. Cl.) E01D 19/12 (2006.01) E01D 2/00 (2006.01) E01D 21/00 (2006.01) (21) 출원번호 10-2011-0036938 (22) 출원일자 2011년04월20일 심사청구일자 2011년04월20일 (65) 공개번호 10-2012-0119156
(19) 대한민국특허청(KR) (12) 등록특허공보(B1) (51) 국제특허분류(Int. Cl.) E01D 19/12 (2006.01) E01D 2/00 (2006.01) E01D 21/00 (2006.01) (21) 출원번호 10-2011-0036938 (22) 출원일자 2011년04월20일 심사청구일자 2011년04월20일 (65) 공개번호 10-2012-0119156
 발간등록번호 11-1470550-000330-01 식품중사용원료진위판별지침서 유전자분석법활용 2013. 12. - 유전자분석법활용 - 발간사 최근부당이익을얻기위하여값싼원료를사용하거나표시사항을허위로기재하는등의불량식품 가짜식품 제조 유통사례가증가하고있는실정입니다. 특히고추양념을혼합한불량고춧가루 다진마늘에양파혼합 홍삼분말에마분말혼합등의사례가언론등을통하여보도되면서식품에대한불안감이있는실정입니다.
발간등록번호 11-1470550-000330-01 식품중사용원료진위판별지침서 유전자분석법활용 2013. 12. - 유전자분석법활용 - 발간사 최근부당이익을얻기위하여값싼원료를사용하거나표시사항을허위로기재하는등의불량식품 가짜식품 제조 유통사례가증가하고있는실정입니다. 특히고추양념을혼합한불량고춧가루 다진마늘에양파혼합 홍삼분말에마분말혼합등의사례가언론등을통하여보도되면서식품에대한불안감이있는실정입니다.
Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정
![Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정](/thumbs/88/116783310.jpg "Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정") Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정보를 RNA로옮겨주는과정 ex) retrovirus 2. DNA Replication ( 유전자복제
Chapter 14 유전정보 (Genetic Information) A + G = C + T 일정 ( 샤가프룰 ] - Watson & Crick(1953년 ) : double helix model of DNA( 이중나선구조 ) 제시 1. 유전정보의전달경로 DNA 상의정보를 RNA로옮겨주는과정 ex) retrovirus 2. DNA Replication ( 유전자복제
붙임2-1. 건강영향 항목의 평가 매뉴얼(협의기관용, '13.12).hwp
.hwp") 환경영향평가서내위생 공중보건항목작성을위한건강영향항목의평가매뉴얼 - 협의기관용 - 2013. 12 환경부환경보건정책관실 - i - - ii - - iii - - iv - - v - - vi - 제 1 장건강영향평가의개요 건강영향평가의정의건강영향평가제도의필요성건강영향평가의목적및기능건강영향평가의원칙건강결정요인 - 1 - - 2 - - 3 - 제 2 장건강영향평가제도의시행방안
환경영향평가서내위생 공중보건항목작성을위한건강영향항목의평가매뉴얼 - 협의기관용 - 2013. 12 환경부환경보건정책관실 - i - - ii - - iii - - iv - - v - - vi - 제 1 장건강영향평가의개요 건강영향평가의정의건강영향평가제도의필요성건강영향평가의목적및기능건강영향평가의원칙건강결정요인 - 1 - - 2 - - 3 - 제 2 장건강영향평가제도의시행방안
1. 세포의구조와구성요소 1.1 세포의구조 표 1.1 원핵세포와진핵세포의비교 원핵세포 핵막없음있음 인없음있음 DNA( 염색체 ) 한개, 히스톤등단백질과결합안됨 분열무사분열유사분열 진핵세포 여러개의염색체로존재. 히스톤등단백질과복합하게결합됨 원형질막보통스테롤이없음보통스테롤
 한개, 히스톤등단백질과결합안됨 분열무사분열유사분열 진핵세포 여러개의염색체로존재. 히스톤등단백질과복합하게결합됨 원형질막보통스테롤이없음보통스테롤") 1. 세포의구조와구성요소 1.1 세포의구조 표 1.1 원핵세포와진핵세포의비교 원핵세포 핵막없음있음 인없음있음 DA( 염색체 ) 한개, 히스톤등단백질과결합안됨 분열무사분열유사분열 진핵세포 여러개의염색체로존재. 히스톤등단백질과복합하게결합됨 원형질막보통스테롤이없음보통스테롤이없음 라이보좀 (ribosome) 세포내소기관 (organelle) 70S(50S+30S)
1. 세포의구조와구성요소 1.1 세포의구조 표 1.1 원핵세포와진핵세포의비교 원핵세포 핵막없음있음 인없음있음 DA( 염색체 ) 한개, 히스톤등단백질과결합안됨 분열무사분열유사분열 진핵세포 여러개의염색체로존재. 히스톤등단백질과복합하게결합됨 원형질막보통스테롤이없음보통스테롤이없음 라이보좀 (ribosome) 세포내소기관 (organelle) 70S(50S+30S)
(01) hwp
 hwp") Journal of Life Science 2013 Vol. 23. No. 2. 157~166 ISSN (Print) 1225-9918 ISSN (Online) 2287-3406 DOI : http://dx.doi.org/10.5352/jls.2013.23.2.157 α μ δ κ 158 생명과학회지 2013, Vol. 23. No. 2 Journal of
Journal of Life Science 2013 Vol. 23. No. 2. 157~166 ISSN (Print) 1225-9918 ISSN (Online) 2287-3406 DOI : http://dx.doi.org/10.5352/jls.2013.23.2.157 α μ δ κ 158 생명과학회지 2013, Vol. 23. No. 2 Journal of
Lehninger Principles of Biochemistry 5/e
 David L. Nelson and Michael M. Cox LEHNINGER PRINCIPLES OF BIOCHEMISTRY Fifth Edition CHAPTER 17 Fatty Acid Catabolism 2008 W. H. Freeman and Company Chap. 17. Fatty acid oxidation 지방산의소화, 동원그리고운반 1. 식이성지방
David L. Nelson and Michael M. Cox LEHNINGER PRINCIPLES OF BIOCHEMISTRY Fifth Edition CHAPTER 17 Fatty Acid Catabolism 2008 W. H. Freeman and Company Chap. 17. Fatty acid oxidation 지방산의소화, 동원그리고운반 1. 식이성지방
3월 온라인 교육
 2013 년 2 분기대리점집체교육 - Protein - Takara Korea Biomedical Protein Workflow Chapter 1: 샘플준비 Chapter 2: Electrophoresis -Precast gel (Lonza/NuSep) -ProSieve EX running buffer -Protein marker Chapter 3: Staining
2013 년 2 분기대리점집체교육 - Protein - Takara Korea Biomedical Protein Workflow Chapter 1: 샘플준비 Chapter 2: Electrophoresis -Precast gel (Lonza/NuSep) -ProSieve EX running buffer -Protein marker Chapter 3: Staining
Cloning
 Takara 와함께하는 Cloning 2014-11-13 다카라코리아바이오메디칼 목차 Cloning 이란? Cloning Flow Chart Cloning DNA / RNA 추출 High Fidelity PCR 제한효소 /ligation/e.coli 형질전환 Clone 확인 이것만은꼭!!! 2 Cloning 이란? Clone 세포나개체의증식에의해서생긴유전적으로동일한세포군
Takara 와함께하는 Cloning 2014-11-13 다카라코리아바이오메디칼 목차 Cloning 이란? Cloning Flow Chart Cloning DNA / RNA 추출 High Fidelity PCR 제한효소 /ligation/e.coli 형질전환 Clone 확인 이것만은꼭!!! 2 Cloning 이란? Clone 세포나개체의증식에의해서생긴유전적으로동일한세포군
- 1 -
 11 1541000-001436-01 수출전략형신수요창출을위한홍마늘및바이오식품개발 (Development of red-garlic and its bio-food for create new market demand and develop of export items) - 1 - - 2 - - 3 - o o - 4 - o o - 5 - o α α - 6 - - 7
11 1541000-001436-01 수출전략형신수요창출을위한홍마늘및바이오식품개발 (Development of red-garlic and its bio-food for create new market demand and develop of export items) - 1 - - 2 - - 3 - o o - 4 - o o - 5 - o α α - 6 - - 7
Chemistry: The Study of Change
 산 - 염기평형및 용해도평형 16.2 공통이온효과 16.3 완충용액 16.4 산 - 염기적정 16.5 산 - 염기지시약 16.6 용해도평형 16.8 공통이온효과및용해도 16.9 ph 및용해도 16.10 착이온평형및용해도 16.7 분별침전에의한이온의분리 Copyright The McGraw-Hill Companies, Inc. Permission required
산 - 염기평형및 용해도평형 16.2 공통이온효과 16.3 완충용액 16.4 산 - 염기적정 16.5 산 - 염기지시약 16.6 용해도평형 16.8 공통이온효과및용해도 16.9 ph 및용해도 16.10 착이온평형및용해도 16.7 분별침전에의한이온의분리 Copyright The McGraw-Hill Companies, Inc. Permission required
- 1 -
 - 1 - - 2 - - 3 - - 4 - 장비구성 : - 5 - - 6 - 치 - 7 - μ - 8 - - 9 - 고체흡착관의안정화방법및기기 (Tube conditioner) - 10 - - 11 - - 12 - - 13 - - 14 - - 15 - - 16 - - 17 - 전기냉각저온농축장치 (TD) GC/FPD - 18 - GC/FID Headspace
- 1 - - 2 - - 3 - - 4 - 장비구성 : - 5 - - 6 - 치 - 7 - μ - 8 - - 9 - 고체흡착관의안정화방법및기기 (Tube conditioner) - 10 - - 11 - - 12 - - 13 - - 14 - - 15 - - 16 - - 17 - 전기냉각저온농축장치 (TD) GC/FPD - 18 - GC/FID Headspace
개최요강
 55 2009. 5. ( ) ( ) < > 1. 1 2. 2. 2. 3 1) 3 2) 3 3) GC-MS 4. 5 1) 5 2) 5 3) ICP-OES 6 3. 7. 7 1) 7 2) 10. 13 1) 1g 13 2),,, 14 3), 15 4),, 15 4. 17 19 < > [ 1] 2 [ 2] ICP-OES 6 [ 3] 13 [ 4] 1g 13 [ 5]
55 2009. 5. ( ) ( ) < > 1. 1 2. 2. 2. 3 1) 3 2) 3 3) GC-MS 4. 5 1) 5 2) 5 3) ICP-OES 6 3. 7. 7 1) 7 2) 10. 13 1) 1g 13 2),,, 14 3), 15 4),, 15 4. 17 19 < > [ 1] 2 [ 2] ICP-OES 6 [ 3] 13 [ 4] 1g 13 [ 5]
<312E20B9DDB5E5BDC320BECFB1E2C7D8BEDF20C7D220C8ADC7D020B9DDC0C0BDC42E687770>
 반드시암기해야할화학반응식 제 1 류위험물 1. 염소산칼륨분해반응식 (400 C) - 2KClO KClO + KCl + O ( 염소산칼륨 ) ( 과염소산칼륨 ) ( 염화칼륨 ) ( 산소 ) 2. 염소산칼륨분해반응식 (540 C~560 C) - 2KClO 2KCl + 3O ( 염소산칼륨 ) ( 염화칼륨 ) ( 산소 ) 3. 염소산칼륨 + 황산 - 6KClO +
반드시암기해야할화학반응식 제 1 류위험물 1. 염소산칼륨분해반응식 (400 C) - 2KClO KClO + KCl + O ( 염소산칼륨 ) ( 과염소산칼륨 ) ( 염화칼륨 ) ( 산소 ) 2. 염소산칼륨분해반응식 (540 C~560 C) - 2KClO 2KCl + 3O ( 염소산칼륨 ) ( 염화칼륨 ) ( 산소 ) 3. 염소산칼륨 + 황산 - 6KClO +
Glutathione Excellose handbook - Purification of GST fusion proteins - Looking for more detail Purification protocol? - Glutathione Excellose Spin Kit
 Glutathione Excellose handbook - Purification of GST fusion proteins - Looking for more detail Purification protocol? - Glutathione Excellose Spin Kit - Glutathione MiniExcellose - Glutathione Excellose
Glutathione Excellose handbook - Purification of GST fusion proteins - Looking for more detail Purification protocol? - Glutathione Excellose Spin Kit - Glutathione MiniExcellose - Glutathione Excellose
PowerPoint 프레젠테이션
 Glutathione Excellose handbook - Purification of GST fusion proteins - Looking for more detail Purification protocol? - Glutathione Excellose Spin Kit - Glutathione MiniExcellose - Glutathione Excellose
Glutathione Excellose handbook - Purification of GST fusion proteins - Looking for more detail Purification protocol? - Glutathione Excellose Spin Kit - Glutathione MiniExcellose - Glutathione Excellose
40.6-(15) fm
 fm") KOREAN J. FOOD SCI. TECHNOL. Vol. 40, No. 6, pp. 696~701 (008) The Korean Society of Food Science and Technology HepG s p l p z w y z w s y z ½ xá x Á xká 1 Á v Áw * w t œw œ, 1 w BK1 ( p pf ), w w t ww
KOREAN J. FOOD SCI. TECHNOL. Vol. 40, No. 6, pp. 696~701 (008) The Korean Society of Food Science and Technology HepG s p l p z w y z w s y z ½ xá x Á xká 1 Á v Áw * w t œw œ, 1 w BK1 ( p pf ), w w t ww
Chapter 14
 5 주 Gycoysis 5.1 Gycoysis의일반적특징 Gycoysis - Gucose가 degradation 과정을거쳐두개의 pyruvate를형성하는과정. - 이때발생하는 free energy는 ATP와 NADH로보존됨. - Gucose 2 Pyruvate - ADP phosphoryation을통하여, ATP 형성 - NAD + 로 hydride ion
5 주 Gycoysis 5.1 Gycoysis의일반적특징 Gycoysis - Gucose가 degradation 과정을거쳐두개의 pyruvate를형성하는과정. - 이때발생하는 free energy는 ATP와 NADH로보존됨. - Gucose 2 Pyruvate - ADP phosphoryation을통하여, ATP 형성 - NAD + 로 hydride ion
생명과학의 이해
 3. 생명현상의 화학적이해 1. 생명체주요원소 2. 생명체주요분자 3. 4. ATP 5. 물 1 1. 생명체주요원소 세포를구성하고있는원소비율원소무게 (%) 1. 산소 (O) 62.00 2. 탄소 (C) 20.00 주성분원소 3. 질소 (N) 10.00 ( 약 95%) 세포를구성하고정상적인기능을위해필요한원소는약 25 종 4. 수소 (H) 3.00 5. 칼슘 (Ca)
3. 생명현상의 화학적이해 1. 생명체주요원소 2. 생명체주요분자 3. 4. ATP 5. 물 1 1. 생명체주요원소 세포를구성하고있는원소비율원소무게 (%) 1. 산소 (O) 62.00 2. 탄소 (C) 20.00 주성분원소 3. 질소 (N) 10.00 ( 약 95%) 세포를구성하고정상적인기능을위해필요한원소는약 25 종 4. 수소 (H) 3.00 5. 칼슘 (Ca)
 MHC T cell Receptor Antigen Antigen-presenting cell Antigen-reactive T cell CD28 B7 Activation (proliferation Cytokine production Cytotoxicity) Antigen-presenting cell Antigen-reactive T cell Dormant State
MHC T cell Receptor Antigen Antigen-presenting cell Antigen-reactive T cell CD28 B7 Activation (proliferation Cytokine production Cytotoxicity) Antigen-presenting cell Antigen-reactive T cell Dormant State
cdna의신장반응이저해된다. 이와같이 AMV 유래 RTase 와 MoMLV 유래 RTase 는모두일장일단을가지나필자는 MoMLV 유래 RTase 를선호한다. 또두효소는최적 ph, 최적염농도등에서도차이가있으므로주의해야한다. 최근 Myers 등은 RTase 활성과 PCR
 RT-PCR 법 II. 역전사효소반응 ( 주 ) 다인바이오연구소 현재분자생물학분야에서 RNA 수준의 gene expression ( 유전자발현 ) 과 cdna (complementary DNA) cloning에널리이용되고있는 RT-PCR (Reverse transcription ploymerase chain reaction) 은생명정보가 DNA에서 RNA로전달된다는분자생물학분야의
RT-PCR 법 II. 역전사효소반응 ( 주 ) 다인바이오연구소 현재분자생물학분야에서 RNA 수준의 gene expression ( 유전자발현 ) 과 cdna (complementary DNA) cloning에널리이용되고있는 RT-PCR (Reverse transcription ploymerase chain reaction) 은생명정보가 DNA에서 RNA로전달된다는분자생물학분야의
Chapter 4. Bacteria (the first microbes)
") - DNA 정제후클로닝을위한효소적절단과연결필요 -> restriction enzyme, ligase 가중요 1. DNA 조작효소범위 - 촉매반응에따라 5 분류 1. Nuclease : 핵산분자의절단, 단편화 2. Ligase : 핵산분자의연결 3. Polymerase : DNA 사슬합성 4. Modifying enzyme : 화학기능기의첨가, 제거 5. Topoisomerase
- DNA 정제후클로닝을위한효소적절단과연결필요 -> restriction enzyme, ligase 가중요 1. DNA 조작효소범위 - 촉매반응에따라 5 분류 1. Nuclease : 핵산분자의절단, 단편화 2. Ligase : 핵산분자의연결 3. Polymerase : DNA 사슬합성 4. Modifying enzyme : 화학기능기의첨가, 제거 5. Topoisomerase
<C8B2BCBABCF62DB8D4B0C5B8AEBFCDB0C7B0AD2E687770>
 먹거리와 건강 황성수 인도주의실천의협의회(humandoctor.org) 황성수 의사님의 글 http://humandoctor.org/technote/main.cgi?board=whang 서적 곰탕이 건강을 말아먹는다 로도 출판되어 있음. lightworker.kr 편집 1. 이 글을 읽는 분들에게 황성수의 먹거리와 건강 을 읽는 모든 분들에게 인사드립니다.
먹거리와 건강 황성수 인도주의실천의협의회(humandoctor.org) 황성수 의사님의 글 http://humandoctor.org/technote/main.cgi?board=whang 서적 곰탕이 건강을 말아먹는다 로도 출판되어 있음. lightworker.kr 편집 1. 이 글을 읽는 분들에게 황성수의 먹거리와 건강 을 읽는 모든 분들에게 인사드립니다.
01 Buffers & Gel Stain Buffers 3 Gel Stain SilverStar Staining Kit 6
 Buffers & Gel Stain Chemicals Buffers & Chemicals Phone: 1588-9788 (ext.4->2) Email: [email protected] 01 Buffers & Gel Stain Buffers 3 Gel Stain SilverStar Staining Kit 6 Buffers Overview
Buffers & Gel Stain Chemicals Buffers & Chemicals Phone: 1588-9788 (ext.4->2) Email: [email protected] 01 Buffers & Gel Stain Buffers 3 Gel Stain SilverStar Staining Kit 6 Buffers Overview
특허청구의범위청구항 1 물을여과하는필터부 ; 상기필터부에물을유동시키는정수관 ; 상기정수관에설치되고, 상기정수관의수류를이용하여전기를발생시키는발전모듈 ; 및상기정수관에배치되고, 상기발전모듈에서발생된전기가공급되고, 상기정수관을따라유동되는정수를전기분해하여살균하는살균모듈 ; 을
 (51) Int. Cl. (19) 대한민국특허청 (KR) (12) 공개특허공보 (A) B01D 35/00 (2006.01) C02F 1/461 (2006.01) C02F 1/467 (2006.01) (21) 출원번호 10-2009-0019000 (22) 출원일자 2009 년 03 월 05 일 심사청구일자 없음 전체청구항수 : 총 4 항 (54) 정수기 (11)
(51) Int. Cl. (19) 대한민국특허청 (KR) (12) 공개특허공보 (A) B01D 35/00 (2006.01) C02F 1/461 (2006.01) C02F 1/467 (2006.01) (21) 출원번호 10-2009-0019000 (22) 출원일자 2009 년 03 월 05 일 심사청구일자 없음 전체청구항수 : 총 4 항 (54) 정수기 (11)
(079-088)Kjbt015.hwp
Kjbt015.hwp") 대한수혈학회지:제18권 제2호, 2007 다중 단일염기 시발체 확장반응을 이용한 ABO 유전자형 검사 현정원 1 장호은 2 허세란 2 송상훈 1 박경운 1,2 송정한 1,2 한규섭 1 = Abstract = 서울대학교 의과대학 검사의학교실 1, 분당서울대학교병원 진단검사의학과 2 ABO Genotyping using a Multiplex Single-base
대한수혈학회지:제18권 제2호, 2007 다중 단일염기 시발체 확장반응을 이용한 ABO 유전자형 검사 현정원 1 장호은 2 허세란 2 송상훈 1 박경운 1,2 송정한 1,2 한규섭 1 = Abstract = 서울대학교 의과대학 검사의학교실 1, 분당서울대학교병원 진단검사의학과 2 ABO Genotyping using a Multiplex Single-base
연구보고서화학물질노출에의한유전자돌연변이 ( 발암 ) 의조기확인 (II) - 실험동물을이용한고감도발암성확인기법의검증 - 임경택 김수진
 의조기확인 (II) - 실험동물을이용한고감도발암성확인기법의검증 - 임경택 김수진") 연구보고서화학물질노출에의한유전자돌연변이 ( 발암 ) 의조기확인 (II) - 실험동물을이용한고감도발암성확인기법의검증 - 임경택 김수진 요약문 i ii 화학물질노출에의한유전자돌연변이 ( 발암 ) 의조기확인 (II) 요약문 iii iv 화학물질노출에의한유전자돌연변이 ( 발암 ) 의조기확인 (II) 요약문 v vi 화학물질노출에의한유전자돌연변이 ( 발암 ) 의조기확인
연구보고서화학물질노출에의한유전자돌연변이 ( 발암 ) 의조기확인 (II) - 실험동물을이용한고감도발암성확인기법의검증 - 임경택 김수진 요약문 i ii 화학물질노출에의한유전자돌연변이 ( 발암 ) 의조기확인 (II) 요약문 iii iv 화학물질노출에의한유전자돌연변이 ( 발암 ) 의조기확인 (II) 요약문 v vi 화학물질노출에의한유전자돌연변이 ( 발암 ) 의조기확인
- 2 -
 터키 / 공통 가이드라인명 GMP Kılavuzu GMP 가이드라인 제정일 상위법 Ÿ Ÿ 제정배경 범위 주요내용 Ÿ Ÿ Ÿ Ÿ Ÿ Ÿ Ÿ 의약품제조시, 제조허가증, 의약품의용도및판매허가요구사항, 안정성, 품질및품질부적합으로인한피해로환자가발생하지않도록제조하기위함임. 화학합성의약품, 생물의약품, 방사성의약품, 임상시험용의약품, 무균의약품, 사람혈액및혈장의약품,
터키 / 공통 가이드라인명 GMP Kılavuzu GMP 가이드라인 제정일 상위법 Ÿ Ÿ 제정배경 범위 주요내용 Ÿ Ÿ Ÿ Ÿ Ÿ Ÿ Ÿ 의약품제조시, 제조허가증, 의약품의용도및판매허가요구사항, 안정성, 품질및품질부적합으로인한피해로환자가발생하지않도록제조하기위함임. 화학합성의약품, 생물의약품, 방사성의약품, 임상시험용의약품, 무균의약품, 사람혈액및혈장의약품,
- 1 -
 - 1 - - 2 - - 3 - - 4 - - 5 - SUMMARY ( 영문요약문 ) - 6 - - 7 - - 8 - - 9 - cavenging - 10 - - 11 - - 12 - - 13 - - 14 - - 15 - - 16 - - 17 - - 18 - - 19 - - 20 - - 21 - - 22 - - 23 - - 24 - - 25 - - 26 -
- 1 - - 2 - - 3 - - 4 - - 5 - SUMMARY ( 영문요약문 ) - 6 - - 7 - - 8 - - 9 - cavenging - 10 - - 11 - - 12 - - 13 - - 14 - - 15 - - 16 - - 17 - - 18 - - 19 - - 20 - - 21 - - 22 - - 23 - - 24 - - 25 - - 26 -
- 2 -
 작품번호 37 Solar material 로쓰일수있는검정색물질의재발견! 출품분야학생부출품부문화학 2009. 5. 13 시 군 학교 ( 소속 ) 학년 ( 직위 ) 성 명 성남시풍생중학교 2 김호기, 이희원 지도교사풍생중학교교사김경원 - 1 - - 2 - - 3 - - 4 - - 5 - - 6 - - 7 - 석탄은주로탄소로구성되어있고, 수소와산소가들어있다. 이밖에질소
작품번호 37 Solar material 로쓰일수있는검정색물질의재발견! 출품분야학생부출품부문화학 2009. 5. 13 시 군 학교 ( 소속 ) 학년 ( 직위 ) 성 명 성남시풍생중학교 2 김호기, 이희원 지도교사풍생중학교교사김경원 - 1 - - 2 - - 3 - - 4 - - 5 - - 6 - - 7 - 석탄은주로탄소로구성되어있고, 수소와산소가들어있다. 이밖에질소
기구명 ph meter volumetric flask Erlenmeyer flask mass cylinder 뷰렛비이커 pipet 저울스탠드 & 클램프 isotonicity 측정기 필요량 500ml짜리 1개, 50ml짜리 5개, 100ml짜리 1개, 250ml짜리 6개
 7. 완충용액 Introduction 일반적으로약산과그짝염기또는약염기와그짝산의혼합물로이루어진용액은외부에서산이나염기를첨가하여도 ph가크게변하지않는다. 이처럼 ph의변화에저항하는용액을완충용액이라한다. 이번실험에서는완충용액을직접제조하여그성질을파악하고완충방정식및완충용액을구해보고자한다. 또한산-염기중화적정곡선을 ph로부터그려보고이로부터산의해리상수도구하여보고자한다. Materials
7. 완충용액 Introduction 일반적으로약산과그짝염기또는약염기와그짝산의혼합물로이루어진용액은외부에서산이나염기를첨가하여도 ph가크게변하지않는다. 이처럼 ph의변화에저항하는용액을완충용액이라한다. 이번실험에서는완충용액을직접제조하여그성질을파악하고완충방정식및완충용액을구해보고자한다. 또한산-염기중화적정곡선을 ph로부터그려보고이로부터산의해리상수도구하여보고자한다. Materials
Microsoft PowerPoint - ch4note
 강의개요 Chapter 4 Reactions in Aqueous Solution ( 수용액중의반응 ) 농도의종류와계산 수용액중의반응 Pb(NO 3 (aq) + 2KI (aq) HCl (aq) + NaOH (aq) Alka-Selter/Water Cu wire/ag(no 3 ) (aq) 침전반응산 - 염기중화반응기체생성반응산화환원반응 용액 (Solution)
강의개요 Chapter 4 Reactions in Aqueous Solution ( 수용액중의반응 ) 농도의종류와계산 수용액중의반응 Pb(NO 3 (aq) + 2KI (aq) HCl (aq) + NaOH (aq) Alka-Selter/Water Cu wire/ag(no 3 ) (aq) 침전반응산 - 염기중화반응기체생성반응산화환원반응 용액 (Solution)
5
 26 3 (2005 9 ) J Korean Oriental Med 2005;26(3):55-65 The Effcects of Psoraleae fructus Extract on Melanin Synthesis of B16 Melanoma Cells Jae-Ho Chung, Hyung-Sik Seo Dept. of Ophthamology, Otolaryngology
26 3 (2005 9 ) J Korean Oriental Med 2005;26(3):55-65 The Effcects of Psoraleae fructus Extract on Melanin Synthesis of B16 Melanoma Cells Jae-Ho Chung, Hyung-Sik Seo Dept. of Ophthamology, Otolaryngology
본 발명은 난연재료 조성물에 관한 것으로, 보다 상세하게는 케이블이나 전선의 시스체로 쓰이는 저발연, 저독성을 가진 열가소성 난연재료 조성물에 관한 것이다. 종래의 선박용 케이블은 그 사용 용도와 장소에 따라 다양한 제품들로 구별된다. 근래 들어 해양 구조물 및 선박에
 (51) Int. Cl. 7 C08L 31/04 (19)대한민국특허청(KR) (12) 등록특허공보(B1) (45) 공고일자 (11) 등록번호 (24) 등록일자 2005년03월08일 10-0473564 2005년02월17일 (21) 출원번호 10-2001-0074372 (65) 공개번호 10-2003-0043291 (22) 출원일자 2001년11월27일 (43)
(51) Int. Cl. 7 C08L 31/04 (19)대한민국특허청(KR) (12) 등록특허공보(B1) (45) 공고일자 (11) 등록번호 (24) 등록일자 2005년03월08일 10-0473564 2005년02월17일 (21) 출원번호 10-2001-0074372 (65) 공개번호 10-2003-0043291 (22) 출원일자 2001년11월27일 (43)
Chapter 11
 15.4 RNA polymerase II core promoter core promotor 지역은전사개시부위에서위로 40bp 아래로 40bp 에해당한다. TATA box TATAXAX consensus sequence located 25-30 bp upstream of the transcription start site initiator element (Inr)
15.4 RNA polymerase II core promoter core promotor 지역은전사개시부위에서위로 40bp 아래로 40bp 에해당한다. TATA box TATAXAX consensus sequence located 25-30 bp upstream of the transcription start site initiator element (Inr)
Microsoft Word doc
 이학석사학위논문 고농도포도당과 Palmitate 에의한 INS-1 베타세포의세포사기작연구 아주대학교대학원 의학과 이연정 고농도포도당과 Palmitate 에의한 INS-1 베타세포의세포사기작연구 지도교수강엽 이논문을이학석사학위논문으로제출함. 2008 년 8 월 아주대학교대학원 의학과 이연정 이연정의이학석사학위논문을인준함. 심사위원장강엽인 심사위원이관우인 심사위원정윤석인
이학석사학위논문 고농도포도당과 Palmitate 에의한 INS-1 베타세포의세포사기작연구 아주대학교대학원 의학과 이연정 고농도포도당과 Palmitate 에의한 INS-1 베타세포의세포사기작연구 지도교수강엽 이논문을이학석사학위논문으로제출함. 2008 년 8 월 아주대학교대학원 의학과 이연정 이연정의이학석사학위논문을인준함. 심사위원장강엽인 심사위원이관우인 심사위원정윤석인
(72) 발명자 이영애 경기도수원시영통구원천동 35 원천주공아파트 102 동 1304 호 쳉퀼링 헹디안인더스트리얼존, 동양시티, 제지앙프로빈스,322118, 중국 우유리앙 헹디안인더스트리얼존, 동양시티, 제지앙프로빈스,322118, 중국 리징 헹디안인더스트리얼존, 동양
 발명자 이영애 경기도수원시영통구원천동 35 원천주공아파트 102 동 1304 호 쳉퀼링 헹디안인더스트리얼존, 동양시티, 제지앙프로빈스,322118, 중국 우유리앙 헹디안인더스트리얼존, 동양시티, 제지앙프로빈스,322118, 중국 리징 헹디안인더스트리얼존, 동양") (51) Int. Cl. (19) 대한민국특허청 (KR) (12) 등록특허공보 (B1) C07C 229/64 (2006.01) C07C 229/38 (2006.01) (21) 출원번호 10-2007-0114660 (22) 출원일자 2007 년 11 월 12 일 심사청구일자 (56) 선행기술조사문헌 과학기술부, 2002 ( 뒷면에계속 ) 2008 년 02 월
(51) Int. Cl. (19) 대한민국특허청 (KR) (12) 등록특허공보 (B1) C07C 229/64 (2006.01) C07C 229/38 (2006.01) (21) 출원번호 10-2007-0114660 (22) 출원일자 2007 년 11 월 12 일 심사청구일자 (56) 선행기술조사문헌 과학기술부, 2002 ( 뒷면에계속 ) 2008 년 02 월
Lehninger Principles of Biochemistry 5/e
 David L. Nelson and Michael M. Cox LEHNINGER PRINCIPLES OF BIOCHEMISTRY Fifth Edition CHAPTER 18 Amino Acid Oxidation and the Production of Urea 2008 W. H. Freeman and Company Dietary proteins Synthesis
David L. Nelson and Michael M. Cox LEHNINGER PRINCIPLES OF BIOCHEMISTRY Fifth Edition CHAPTER 18 Amino Acid Oxidation and the Production of Urea 2008 W. H. Freeman and Company Dietary proteins Synthesis
..........
 통권 제101호(2007년 10월호) 특허가족 건강길라잡이(비만 시리즈4) 특허가족 건강길라잡이 (비만 시리즈 4) 변리사 김 일 성 킴스국제특허법률사무소 2. 비만과 영양소 4. 무기질(Minerals) 무기질은 단백질, 당질, 지질들과 달리 에너지를 낼 수 없고, 비타민류와 달리 조리과정에서 파괴되지 않는다. 그러나 물에 용해되므로 조리 중의 용액을 버리
통권 제101호(2007년 10월호) 특허가족 건강길라잡이(비만 시리즈4) 특허가족 건강길라잡이 (비만 시리즈 4) 변리사 김 일 성 킴스국제특허법률사무소 2. 비만과 영양소 4. 무기질(Minerals) 무기질은 단백질, 당질, 지질들과 달리 에너지를 낼 수 없고, 비타민류와 달리 조리과정에서 파괴되지 않는다. 그러나 물에 용해되므로 조리 중의 용액을 버리
Microsoft Word - 04-김택중.doc
 J. Exp. Biomed. Sci. 2010, 16(2): 91~95 Anti-inflammatory Properties of Meso-dihydroguaiaretic Acid in Lipopolysaccharide-induced Macrophage Yong-Jae Kim *, Yeo-Jin Kang * and Tack-Joong Kim Division of
J. Exp. Biomed. Sci. 2010, 16(2): 91~95 Anti-inflammatory Properties of Meso-dihydroguaiaretic Acid in Lipopolysaccharide-induced Macrophage Yong-Jae Kim *, Yeo-Jin Kang * and Tack-Joong Kim Division of
슬라이드 1
 LC-MS 단백질분석을위한 SDS-PAGE staining protocol 및주의할점 Proteomics Core Facility 2017.12.28 Contents LC-MS 분석의뢰용 protein SDS-PAGE staining 시주의할점 Coomassie Blue R-250 을이용한 gel staining (general protocol) Coomassie
LC-MS 단백질분석을위한 SDS-PAGE staining protocol 및주의할점 Proteomics Core Facility 2017.12.28 Contents LC-MS 분석의뢰용 protein SDS-PAGE staining 시주의할점 Coomassie Blue R-250 을이용한 gel staining (general protocol) Coomassie
PowerPoint Presentation
 Interchapeter B : 핵산생물공학기술 (part A) Recombinant DNA techniques - 전기영동 - 제한효소 - 클로닝 - 대장균을이용한단백질대량생산 Agarose mesh DNA 분리기술 : Agarose Gel Electrophoresis ( 수평형 ) _ + DNA is negatively charged from the phosphate
Interchapeter B : 핵산생물공학기술 (part A) Recombinant DNA techniques - 전기영동 - 제한효소 - 클로닝 - 대장균을이용한단백질대량생산 Agarose mesh DNA 분리기술 : Agarose Gel Electrophoresis ( 수평형 ) _ + DNA is negatively charged from the phosphate
(72) 발명자 도리야마 킨야 일본 미야기켄 센다이시 아오바쿠 가타히라 2쵸메 1반 1고 고쿠리츠 다이가쿠 호진 도호쿠 다이가쿠 내 오가와 도모히사 일본 미야기켄 센다이시 아오바쿠 가타히라 2쵸메 1반 1고 고쿠리츠 다이가쿠 호진 도호쿠 다이가쿠 내 무라모토 고지 일본
 발명자 도리야마 킨야 일본 미야기켄 센다이시 아오바쿠 가타히라 2쵸메 1반 1고 고쿠리츠 다이가쿠 호진 도호쿠 다이가쿠 내 오가와 도모히사 일본 미야기켄 센다이시 아오바쿠 가타히라 2쵸메 1반 1고 고쿠리츠 다이가쿠 호진 도호쿠 다이가쿠 내 무라모토 고지 일본") (19) 대한민국특허청(KR) (12) 공개특허공보(A) (51) 국제특허분류(Int. Cl.) A01H 5/00 (2006.01) C12N 15/82 (2006.01) C12N 15/29 (2006.01) C07K 14/42 (2006.01) (21) 출원번호 10-2012-7012238 (22) 출원일자(국제) 2010년10월14일 심사청구일자 없음 (85)
(19) 대한민국특허청(KR) (12) 공개특허공보(A) (51) 국제특허분류(Int. Cl.) A01H 5/00 (2006.01) C12N 15/82 (2006.01) C12N 15/29 (2006.01) C07K 14/42 (2006.01) (21) 출원번호 10-2012-7012238 (22) 출원일자(국제) 2010년10월14일 심사청구일자 없음 (85)
미생물분류는형태적특징, 생리 생화학적성질과상태, 화학분류학적성질과상태등을이용하여구분하는것이일반적이지만, 이와같은방법을이용하면많은시간을필요로한다. 또한분류가힘든경우나, 정확하지못한결과를얻는경우도있다. 최근미생물분류에도분자생물학적인방법을이용하여, 미생물이가지고있는 DNA를
 Code RR180A - 연구용 - Bacterial 16S rdna PCR Kit 사용설명서 v201011da 미생물분류는형태적특징, 생리 생화학적성질과상태, 화학분류학적성질과상태등을이용하여구분하는것이일반적이지만, 이와같은방법을이용하면많은시간을필요로한다. 또한분류가힘든경우나, 정확하지못한결과를얻는경우도있다. 최근미생물분류에도분자생물학적인방법을이용하여, 미생물이가지고있는
Code RR180A - 연구용 - Bacterial 16S rdna PCR Kit 사용설명서 v201011da 미생물분류는형태적특징, 생리 생화학적성질과상태, 화학분류학적성질과상태등을이용하여구분하는것이일반적이지만, 이와같은방법을이용하면많은시간을필요로한다. 또한분류가힘든경우나, 정확하지못한결과를얻는경우도있다. 최근미생물분류에도분자생물학적인방법을이용하여, 미생물이가지고있는
<4D F736F F F696E74202D20322DC0AFC0FCC0DAB9DFC7F6C0C7C1B6C0FD205BC8A3C8AF20B8F0B5E55D>
 주제 2. 유전자발현의조절 생명과학 11 장앞부분 Figure 06.01: The minimum gene number required for any type of organism increases with its complexity. 1 인간게놈 DNA 중 : 유전자 ( 엑손 + 인트론 ): 25% 엑손 : 1%, 인트론 : 24% 나머지는유전자가아닌 DNA
주제 2. 유전자발현의조절 생명과학 11 장앞부분 Figure 06.01: The minimum gene number required for any type of organism increases with its complexity. 1 인간게놈 DNA 중 : 유전자 ( 엑손 + 인트론 ): 25% 엑손 : 1%, 인트론 : 24% 나머지는유전자가아닌 DNA
1. A B C 4. ABC B C A B A B C A C AB BC ABC. ABC C + A + B C A B A B C A B C B A C B C A C A B C B A 5. AB xy pqr x B xy p -y AB. A. A. B. TV B. C. AB
 018 학년도대학수학능력시험문제및정답 1. A B C 4. ABC B C A B A B C A C AB BC ABC. ABC C + A + B C A B A B C A B C B A C B C A C A B C B A 5. AB xy pqr x B xy p -y AB. A. A. B. TV B. C. ABC lålılç lå
018 학년도대학수학능력시험문제및정답 1. A B C 4. ABC B C A B A B C A C AB BC ABC. ABC C + A + B C A B A B C A B C B A C B C A C A B C B A 5. AB xy pqr x B xy p -y AB. A. A. B. TV B. C. ABC lålılç lå
Product Description Apoptosis 혹은 necrosis등에의하여죽거나손상된세포에서방출되는 Lactate dehydrogenase(ldh) 의양을고감도로측정함으로써 cytotoxicity/cytolysis를간단하게측정할수있는 kit 입니다. Cytot
 의양을고감도로측정함으로써 cytotoxicity/cytolysis를간단하게측정할수있는 kit 입니다. Cytot") EZ-LDH Cell Cytotoxicity Assay Kit Cat. No. DG-LDH500 DG-LDH1000 FOR RESEARCH USE ONLY. NOT FOR USE IN DIAGNOSTIC PROCEDURES. 1 ( 주 ) 대일랩서비스부설두젠생명공학연구소 Product Description Apoptosis 혹은 necrosis등에의하여죽거나손상된세포에서방출되는
EZ-LDH Cell Cytotoxicity Assay Kit Cat. No. DG-LDH500 DG-LDH1000 FOR RESEARCH USE ONLY. NOT FOR USE IN DIAGNOSTIC PROCEDURES. 1 ( 주 ) 대일랩서비스부설두젠생명공학연구소 Product Description Apoptosis 혹은 necrosis등에의하여죽거나손상된세포에서방출되는
학술원논문집 ( 자연과학편 ) 제 50 집 2 호 (2011) 콩의식품적의의및생산수급과식용콩의자급향상 李弘䄷 * 李英豪 ** 李錫河 *** * Significance of Soybean as Food and Strategies for Self Suffici
 제 50 집 2 호 (2011) 콩의식품적의의및생산수급과식용콩의자급향상 李弘䄷 * 李英豪 ** 李錫河 *** * Significance of Soybean as Food and Strategies for Self Suffici") 학술원논문집 ( 자연과학편 ) 제 50 집 2 호 (2011) 97-137 콩의식품적의의및생산수급과식용콩의자급향상 李弘䄷 * 李英豪 ** 李錫河 *** * Significance of Soybean as Food and Strategies for Self Sufficiency Improvement Hong Suk Lee*, Yeong-Ho Lee**, and
학술원논문집 ( 자연과학편 ) 제 50 집 2 호 (2011) 97-137 콩의식품적의의및생산수급과식용콩의자급향상 李弘䄷 * 李英豪 ** 李錫河 *** * Significance of Soybean as Food and Strategies for Self Sufficiency Improvement Hong Suk Lee*, Yeong-Ho Lee**, and
어린이기호식품1회제공량표시-결과보고서-줄임.hwp
 우리는소비자와함께행복한세상을만든다 어린이기호식품 1 회제공량표시조사보고서 2010. 11 식품미생물팀 차례 Ⅰ. 조사개요 1 1. 추진배경 1 2. 조사기간및조사내용 2 Ⅱ. 조사결과 2 1. 어린이기호식품 2 2. 가공식품의영양표시제도 4 3. 1회제공량표시 9 4. 영양성분앞면표시제 12 5. 신호등표시제 14 6. 고열량 저영양식품 16 7. 어린이기호식품품질인증
우리는소비자와함께행복한세상을만든다 어린이기호식품 1 회제공량표시조사보고서 2010. 11 식품미생물팀 차례 Ⅰ. 조사개요 1 1. 추진배경 1 2. 조사기간및조사내용 2 Ⅱ. 조사결과 2 1. 어린이기호식품 2 2. 가공식품의영양표시제도 4 3. 1회제공량표시 9 4. 영양성분앞면표시제 12 5. 신호등표시제 14 6. 고열량 저영양식품 16 7. 어린이기호식품품질인증
PowerPoint 프레젠테이션
 2016-10-25 KU Microbial Biotechnology Laboratory 1 KUmbl 경북대학교미생물공학연구실 7 장. 아미노산발효 2016-10-25 KU Microbial Biotechnology Laboratory 2 7-0. 아미노산발효의종류 * 아미노산발효의종류 1. 직접발효법 : 미생물을이용하여기질로부터발효에의해생산 1) 야생균주에의한방법
2016-10-25 KU Microbial Biotechnology Laboratory 1 KUmbl 경북대학교미생물공학연구실 7 장. 아미노산발효 2016-10-25 KU Microbial Biotechnology Laboratory 2 7-0. 아미노산발효의종류 * 아미노산발효의종류 1. 직접발효법 : 미생물을이용하여기질로부터발효에의해생산 1) 야생균주에의한방법
(72) 발명자 장종산 대전 중구 수침로 138, 103동 204호 (태평동, 유등 마을쌍용아파트) 박용기 대전 유성구 어은로 57, 119동 302호 (어은동, 한 빛아파트) 황동원 경기 안양시 만안구 양화로147번길 7, 102동 403호 (박달동, 박달동동원베네스
 발명자 장종산 대전 중구 수침로 138, 103동 204호 (태평동, 유등 마을쌍용아파트) 박용기 대전 유성구 어은로 57, 119동 302호 (어은동, 한 빛아파트) 황동원 경기 안양시 만안구 양화로147번길 7, 102동 403호 (박달동, 박달동동원베네스") (19) 대한민국특허청(KR) (12) 공개특허공보(A) (11) 공개번호 10-2015-0069268 (43) 공개일자 2015년06월23일 (51) 국제특허분류(Int. Cl.) B01J 20/10 (2006.01) B01D 53/62 (2006.01) B01J 20/282 (2006.01) (21) 출원번호 10-2013-0155502 (22) 출원일자
(19) 대한민국특허청(KR) (12) 공개특허공보(A) (11) 공개번호 10-2015-0069268 (43) 공개일자 2015년06월23일 (51) 국제특허분류(Int. Cl.) B01J 20/10 (2006.01) B01D 53/62 (2006.01) B01J 20/282 (2006.01) (21) 출원번호 10-2013-0155502 (22) 출원일자